U V W X Y Z - Nutrition
UFC
Urine
Vitamin A
Vitamin B3 Niacin
Vitamin B9 Folate/Folic Acid
Vitamin B12
Vitamin C
Vitamin D
Vitamin E
Vitamin K
Volatile Fatty Acids (VFAs - also SCFAs)
Water
Wheat bran - see Bran
Wheatfeed
Whey protein
Willow / white willow / salicin/salicylic acid
Wood chewing
Yea-sacc
Zeolite
Zinc
Urine
Vitamin A
Vitamin B3 Niacin
Vitamin B9 Folate/Folic Acid
Vitamin B12
Vitamin C
Vitamin D
Vitamin E
Vitamin K
Volatile Fatty Acids (VFAs - also SCFAs)
Water
Wheat bran - see Bran
Wheatfeed
Whey protein
Willow / white willow / salicin/salicylic acid
Wood chewing
Yea-sacc
Zeolite
Zinc
UFC
"The French Net Energy system (primarily developed by the Institut National de la Recherche Agronomique [INRA]) was developed to allow for the differences in utilization of the metabolizable energy available from different feeds, depending on the proportion of the end products of digestion produced and the biochemical pathways used by these end products to produce mechanical energy (as outlined above). It uses the horse feed unit (HFU) or in French, l’unite fouragire cheval (UFC). The UFC corresponds to the net energy value (2250 kcal) of one kg standard barley (87% DM) in a horse at maintenance. The UFC value of a particular feed is calculated by dividing its NE content in kcal by that of barley, i.e. 2250.
The NE UFC/kg DM for corn, barley, oats, maize silage, hay and straw are 1.35, 1.16, 1.01, 0.88, 0.53 and 0.28, respectively. Comparing this system with the DE/ME system, the energy value of straw for example has been reported to be 41% that of barley using DE values, 37% that of barley using ME values and 29% that of barley using NE. Using this as an extreme example, if a 500 kg horse was being fed 3 kg barley and 7 kg hay and one wanted to replace the barley, in energy terms, with straw : · Using DE values one would calculate having to feed about 7.5 kgs of straw. · Using NE values (to get the same useable energy ) one would calculate having to feed about 10.5 kgs of straw. The discrepancy between the actual energy value to the horse for different feeds as calculated by these various methods increases as the cell wall contents increase. So if you fed a diet which contained certain high fiber feedstuffs according to its DE value you would tend to overestimate the actual useable energy available from that diet and the horse might either lose weight or lack sufficient energy. For the purpose of this paper the NE value for oats (as fed) according to the French Net Energy system is taken to be ~2.0 MCal/kg and for hay ~1.0 – 1.25 Mcal/kg."
COMPARISON OF THE DIGESTIBLE ENERGY (DE) AND NET ENERGY (NE) SYSTEMS FOR THE HORSE PAT A. HARRIS
Valeur énergétique des aliments - van den Boorn horses
"Unité Fourragère de Cheval (UFC) was converted to ME using INRA’s ME value of 1 kg of barley, 12.05 MJ, and converting metabolic body weight (MBW) to body weight (BW) on the basis of a 500-kg horse."
Mark Ebert, Meriel J S Moore-Colyer
The energy requirements of performance horses in training
Translational Animal Science, Volume 4, Issue 2, April 2020, Pages 569–588, https://doi.org/10.1093/tas/txaa032
"The French Net Energy system (primarily developed by the Institut National de la Recherche Agronomique [INRA]) was developed to allow for the differences in utilization of the metabolizable energy available from different feeds, depending on the proportion of the end products of digestion produced and the biochemical pathways used by these end products to produce mechanical energy (as outlined above). It uses the horse feed unit (HFU) or in French, l’unite fouragire cheval (UFC). The UFC corresponds to the net energy value (2250 kcal) of one kg standard barley (87% DM) in a horse at maintenance. The UFC value of a particular feed is calculated by dividing its NE content in kcal by that of barley, i.e. 2250.
The NE UFC/kg DM for corn, barley, oats, maize silage, hay and straw are 1.35, 1.16, 1.01, 0.88, 0.53 and 0.28, respectively. Comparing this system with the DE/ME system, the energy value of straw for example has been reported to be 41% that of barley using DE values, 37% that of barley using ME values and 29% that of barley using NE. Using this as an extreme example, if a 500 kg horse was being fed 3 kg barley and 7 kg hay and one wanted to replace the barley, in energy terms, with straw : · Using DE values one would calculate having to feed about 7.5 kgs of straw. · Using NE values (to get the same useable energy ) one would calculate having to feed about 10.5 kgs of straw. The discrepancy between the actual energy value to the horse for different feeds as calculated by these various methods increases as the cell wall contents increase. So if you fed a diet which contained certain high fiber feedstuffs according to its DE value you would tend to overestimate the actual useable energy available from that diet and the horse might either lose weight or lack sufficient energy. For the purpose of this paper the NE value for oats (as fed) according to the French Net Energy system is taken to be ~2.0 MCal/kg and for hay ~1.0 – 1.25 Mcal/kg."
COMPARISON OF THE DIGESTIBLE ENERGY (DE) AND NET ENERGY (NE) SYSTEMS FOR THE HORSE PAT A. HARRIS
Valeur énergétique des aliments - van den Boorn horses
"Unité Fourragère de Cheval (UFC) was converted to ME using INRA’s ME value of 1 kg of barley, 12.05 MJ, and converting metabolic body weight (MBW) to body weight (BW) on the basis of a 500-kg horse."
Mark Ebert, Meriel J S Moore-Colyer
The energy requirements of performance horses in training
Translational Animal Science, Volume 4, Issue 2, April 2020, Pages 569–588, https://doi.org/10.1093/tas/txaa032
Urine
Horse urine varies considerably in colour from almost colourless to golden or brownish, and also in density, turbidity and mucinous content.
Dipsticks can be used to measure pH (urine samples should be taken mid-stream).
Normal urine pH for adult horses is 7.5-8.5, for foals 5.5-8.0.
Urine pH reflects diet. Horses eating mostly pasture and forage will normally have alkaline urine, horses eating a cereal based diet may have slightly acidic urine. Urine will usually contain calcium carbonate crystals.
Proteinuria may be seen with inflammation or renal tubular pathology.
Horses with PPID may have glucosuria.
Urinalysis - Rossdales Laboratories
Horse urine varies considerably in colour from almost colourless to golden or brownish, and also in density, turbidity and mucinous content.
Dipsticks can be used to measure pH (urine samples should be taken mid-stream).
Normal urine pH for adult horses is 7.5-8.5, for foals 5.5-8.0.
Urine pH reflects diet. Horses eating mostly pasture and forage will normally have alkaline urine, horses eating a cereal based diet may have slightly acidic urine. Urine will usually contain calcium carbonate crystals.
Proteinuria may be seen with inflammation or renal tubular pathology.
Horses with PPID may have glucosuria.
Urinalysis - Rossdales Laboratories
Vitamins
Vitamins are carbon-containing micronutrients produced by living cells and required in tiny concentrations.
Vitamins are water (Bs and C) or fat (A, D, E and K) soluble. Fat soluble vitamins are absorbed in association with fat in the diet, stored in lipid tissues in the body and excreted in the dung. Water soluble vitamins serve as coenzymes (catalysts) for many metabolic reactions, are not stored long-term in the body, and are excreted in urine.
Vitamins A and E must come from the diet. Vitamins C and D can by synthesized in the horse. All B vitamins and vitamin K are produced by the horse's intestinal microbiome. However, there may be a requirement for B1 in the diet in addition to intestinal production.
There is little evidence that supplementation of vitamins above established requirements gives any benefit to a horse's health or performance.
Horses grazing pasture will generally eat sufficient vitamin A and E, synthesize sufficient vitamin D from sunlight, and get enough B vitamins and vitamin K from gut microbes, and vitamin C from liver synthesize. However, horses that do not have fresh grass as the main part of their diet may need supplementation with vitamins (in a balancer or mineral and vitamin mix).
Vitamins are carbon-containing micronutrients produced by living cells and required in tiny concentrations.
Vitamins are water (Bs and C) or fat (A, D, E and K) soluble. Fat soluble vitamins are absorbed in association with fat in the diet, stored in lipid tissues in the body and excreted in the dung. Water soluble vitamins serve as coenzymes (catalysts) for many metabolic reactions, are not stored long-term in the body, and are excreted in urine.
Vitamins A and E must come from the diet. Vitamins C and D can by synthesized in the horse. All B vitamins and vitamin K are produced by the horse's intestinal microbiome. However, there may be a requirement for B1 in the diet in addition to intestinal production.
There is little evidence that supplementation of vitamins above established requirements gives any benefit to a horse's health or performance.
Horses grazing pasture will generally eat sufficient vitamin A and E, synthesize sufficient vitamin D from sunlight, and get enough B vitamins and vitamin K from gut microbes, and vitamin C from liver synthesize. However, horses that do not have fresh grass as the main part of their diet may need supplementation with vitamins (in a balancer or mineral and vitamin mix).
Vitamin A
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
α-Carotene, β-Carotene, β-Cryptoxanthin, Lycopene, Lutein, and Zeaxanthin - Linus Pauling Institute
Pass the Carrots: Beta-Carotene for Horses - KER Feb 2011
Vitamin A - human research/information
Vitamin A Fact Sheet for Health Professionals NIH 2020
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
α-Carotene, β-Carotene, β-Cryptoxanthin, Lycopene, Lutein, and Zeaxanthin - Linus Pauling Institute
Pass the Carrots: Beta-Carotene for Horses - KER Feb 2011
Vitamin A - human research/information
Vitamin A Fact Sheet for Health Professionals NIH 2020
Vitamin B1 Thiamine
Vitamin B1 plays a central role in the release of energy from carbohydrates (e.g. the conversion of pyruvate to acetyl coenzyme A), is involved in DNA and RNA production and nerve function. The active form is the coenzyme thiamine pyrophosphate (TPP).
Vitamin B1 plays a central role in the release of energy from carbohydrates (e.g. the conversion of pyruvate to acetyl coenzyme A), is involved in DNA and RNA production and nerve function. The active form is the coenzyme thiamine pyrophosphate (TPP).
Vitamin B3 Niacin
Niacin refers to nicotinic acid and the amide nicotinamide. Nicotinic acid in the diet is converted to nicotinamide and absorbed by intestinal mucosa. The active forms are coenzymes nicotinamide adenine dinucleotide (NAD) (essential for oxidative phosphorylation in energy metabolism) and nicotinamide adenine dinucleotide phosphate (NADP). Niacin is important for mitochondrial respiration and in the metabolism of carbohydrates, fats and amino acids. Niacin is not stored in the body and is rapidly excreted in urine.
Niacin has been used in humans for the treatment of dyslipidaemias as it decreases triglycerides and LDL cholesterol and increases HDL cholesterol.
Niacin is produced by microbes in the horse's digestive tract, and is present in all living tissues. Niacin is found in leafy forages (24-42 mg/kg DM), but although niacin is found in cereal grains (16-94 mg/kg DM) it is in an unavailable form. Niacin decreases at a rate of around 5% per month in stored feed, so year old hay will still contain around 50% of its original niacin concentration. Niacin can be synthesized from the amino acid tryptophan in the horse's liver (but high levels of the amino acid leucine found in cereal grains may decrease tryptophan conversion to niacin).
In conclusion, enough niacin is found in normal horse forages and feeds, along with microbial and liver synthesis, that dietary supplementation of niacin is not required for horses.
Niacin and insulin
There is some research in other species (including humans) that suggests that long-term supplementation of niacin causes insulin resistance - it may be sensible not to oversupplement niacin for horses with insulin dysregulation.
Vitamin B3 - HorseDVM
"research suggests prolonged niacin supplementation increases insulin resistance because it hinders the ability of insulin to suppress glucose synthesis in the liver."
Sun W, Zhai M, Zhou Q, Qian C, Jiang C.
Effects of B Vitamins Overload on Plasma Insulin Level and Hydrogen Peroxide Generation in Rats.
Chin J Physiol. 2017 Aug 31;60(4):207-214. doi: 10.4077/CJP.2017.BAF469. PMID: 28847140.
Excess vitamin B1 and B3 caused oxidative stress and insulin resistance, and cumulative administration of nicotinic acid or nicotinamide impaired glucose metabolism in rats.
Heemskerk MM, van den Berg SA, Pronk AC, van Klinken JB, Boon MR, Havekes LM, Rensen PC, van Dijk KW, van Harmelen V.
Long-term niacin treatment induces insulin resistance and adrenergic responsiveness in adipocytes by adaptive downregulation of phosphodiesterase 3B.
Am J Physiol Endocrinol Metab. 2014 Apr 1;306(7):E808-13. doi: 10.1152/ajpendo.00641.2013. Epub 2014 Jan 28. PMID: 24473440; PMCID: PMC3962609.
"In conclusion, long-term niacin treatment resulted in insulin resistance and increased β-adrenergic responsiveness".
Niacin refers to nicotinic acid and the amide nicotinamide. Nicotinic acid in the diet is converted to nicotinamide and absorbed by intestinal mucosa. The active forms are coenzymes nicotinamide adenine dinucleotide (NAD) (essential for oxidative phosphorylation in energy metabolism) and nicotinamide adenine dinucleotide phosphate (NADP). Niacin is important for mitochondrial respiration and in the metabolism of carbohydrates, fats and amino acids. Niacin is not stored in the body and is rapidly excreted in urine.
Niacin has been used in humans for the treatment of dyslipidaemias as it decreases triglycerides and LDL cholesterol and increases HDL cholesterol.
Niacin is produced by microbes in the horse's digestive tract, and is present in all living tissues. Niacin is found in leafy forages (24-42 mg/kg DM), but although niacin is found in cereal grains (16-94 mg/kg DM) it is in an unavailable form. Niacin decreases at a rate of around 5% per month in stored feed, so year old hay will still contain around 50% of its original niacin concentration. Niacin can be synthesized from the amino acid tryptophan in the horse's liver (but high levels of the amino acid leucine found in cereal grains may decrease tryptophan conversion to niacin).
In conclusion, enough niacin is found in normal horse forages and feeds, along with microbial and liver synthesis, that dietary supplementation of niacin is not required for horses.
Niacin and insulin
There is some research in other species (including humans) that suggests that long-term supplementation of niacin causes insulin resistance - it may be sensible not to oversupplement niacin for horses with insulin dysregulation.
Vitamin B3 - HorseDVM
"research suggests prolonged niacin supplementation increases insulin resistance because it hinders the ability of insulin to suppress glucose synthesis in the liver."
Sun W, Zhai M, Zhou Q, Qian C, Jiang C.
Effects of B Vitamins Overload on Plasma Insulin Level and Hydrogen Peroxide Generation in Rats.
Chin J Physiol. 2017 Aug 31;60(4):207-214. doi: 10.4077/CJP.2017.BAF469. PMID: 28847140.
Excess vitamin B1 and B3 caused oxidative stress and insulin resistance, and cumulative administration of nicotinic acid or nicotinamide impaired glucose metabolism in rats.
Heemskerk MM, van den Berg SA, Pronk AC, van Klinken JB, Boon MR, Havekes LM, Rensen PC, van Dijk KW, van Harmelen V.
Long-term niacin treatment induces insulin resistance and adrenergic responsiveness in adipocytes by adaptive downregulation of phosphodiesterase 3B.
Am J Physiol Endocrinol Metab. 2014 Apr 1;306(7):E808-13. doi: 10.1152/ajpendo.00641.2013. Epub 2014 Jan 28. PMID: 24473440; PMCID: PMC3962609.
"In conclusion, long-term niacin treatment resulted in insulin resistance and increased β-adrenergic responsiveness".
Vitamin B5 Pantothenic acid
Vitamin B5 plays a role in the oxidation of fatty acids and carbohydrates. Coenzyme A is synthesized from vitamin B5 and is involved in the synthesis of amino acids, fatty acids, cholesterol, phospholipids, steroid hormones, neurotransmitters and antibodies.
Vitamin B5 plays a role in the oxidation of fatty acids and carbohydrates. Coenzyme A is synthesized from vitamin B5 and is involved in the synthesis of amino acids, fatty acids, cholesterol, phospholipids, steroid hormones, neurotransmitters and antibodies.
Vitamin B9 Folate/Folic Acid
Roberts MC.
Serum and red cell folate and serum vitamin B12 levels in horses.
Aust Vet J. 1983 Apr;60(4):106-11. doi: 10.1111/j.1751-0813.1983.tb05906.x.
Roberts MC.
Serum and red cell folate and serum vitamin B12 levels in horses.
Aust Vet J. 1983 Apr;60(4):106-11. doi: 10.1111/j.1751-0813.1983.tb05906.x.
Vitamin B12
Vitamin B12 is a water-soluble vitamin and cannot be stored in the body (in significant amounts).
There is no dietary requirement for vitamin B12 above that supplied by intestinal synthesis.
Horses (only) need cobalt in their diet so that their gut bacteria can produce vitamin B12.
There appears to be enough cobalt in forage and typical horse diets to meet cobalt and therefore vitamin B12 requirements.
A healthy hindgut microbiome is important for vitamin B12 production. In cattle, a high forage diet was linked to 3 x more vitamin B12 production than a high starch diet.
Supplementation with B12 might be considered following antibiotics or NSAIDs, diet changes, GI illness e.g. diarrhoea, high-intensity training or stressful events, general ill health or reduced appetite, for young or very old horses - any time the hind gut microbiome might be adversely affected.
Neither deficiency nor toxicity of B12 has been described in horses.
Vitamin B12 is also called cobalamin. B12 is synthesised naturally by microorganisms during hind gut fermentation, and absorbed from the caecum and colon. Horses are thought to have no requirement for B12 supplementation. Plants do not contain B12. Synthesis of B12 requires cobalt.
Cobalt and B12 are important for:
energy metabolism, e.g. the formation of glucose from volatile fatty acids, protein synthesis and fat metabolism,
the synthesis of myelin which coats nerves,
the maturation of red blood cells in bone marrow.
Caple et al. 1982 examined several hundred horses and reported plasma vitamin B12 concentrations between 1.8 and 7.3 micrograms/L. Intramuscular injections of vitamin B12 were rapidly cleared from the plasma, and large amounts excreted in the faeces via the bile when vitamin B12 was administered intravenously to foals.
There is very little research about cobalt in horses. Systemic absorption was demonstrated from the hindgut after administration of cyanocobalamin cobalt into the caecum (Stillions 1971). Cobalt is absorbed as part of the cobalamin complex formed through bacterial synthesis, and a constant replenishment of cobalt is required for cobalamin synthesis. "Microbial synthesis of cobalamin by the equine intestinal community occurs and obviously appears to provide sufficient cobalamin for the horse (Salminen 1975)" (Source: Equine Applied and Clinical Nutrition 2013). 0.1 mg/kg DM diet is considered adequate (McDowell 2003). Sources of cobalt: grass pasture 0.5-0.2 mg/kg DM, alfalfa 0.3 mg/kg DM, yeast is also a good source of cobalt. Straw is low in cobalt.
Source: Equine Applied and Clinical Nutrition 2013, NRC Nutrient Requirements of Horses 2007.
Vitamin B12 - Do Horses Need It In Their Diet? Nerida Richards
Can Vitamin B12 Improve a Horse's Appetite? Clair Thunes July 2018 www.thehorse.com
The Story of B Vitamins in Horse Nutrition - KER June 2012
Hillyer LL, Ridd Z, Fenwick S, Hincks P, Paine SW.
Pharmacokinetics of inorganic cobalt and a vitamin B12 supplement in the Thoroughbred horse: Differentiating cobalt abuse from supplementation.
Equine Vet J. 2018 May;50(3):343-349. doi: 10.1111/evj.12774. Epub 2017 Nov 12. PMID: 29053883.
Wenzel R, Major D, Hesp K, Doble P.
Determination of vitamin B12 in equine urine by liquid chromatography - inductively coupled plasma - mass spectrometry.
J Trace Elem Med Biol. 2018 Dec;50:634-639. doi: 10.1016/j.jtemb.2018.05.005. Epub 2018 May 9. PMID: 29773468.
"a horse can excrete a significant proportion of a commercially available vitamin B12 injection in urine without metabolic modification."
Roberts MC.
Serum and red cell folate and serum vitamin B12 levels in horses.
Aust Vet J. 1983 Apr;60(4):106-11. doi: 10.1111/j.1751-0813.1983.tb05906.x.
Vitamin B12 injections
Vitbee is an injection for horses and cattle containing 0.100% w/v cyanocobalamin.
"Indications for use, specifying the target species: As an aid in raising Vitamin B12 levels in cattle and horses, calves and foals. For use in the treatment of Vitamin B12 deficiency, and for poor growth rates and general unthriftiness in young animals when associated with the above deficiency." "Following treatment, the vitamin B12 status of herbivores should be maintained by dietary supplementation with cobalt."
Butasal is an injection for horses, cattle, dogs and cats containing phosphorus as Butafosfan and 0.05 mg cyanocobalamin (vitamin B12) per ml.
"Cyanocobalamin is a co-enzyme in the biosynthesis of glucose from propionate. Further it serves as a co-factor to enzymes important in fatty acid synthesis and is important for maintenance of normal haemopoiesis, protection of the liver, and maintenance of muscle tissue, healthy skin, brain and pancreatic metabolism. It belongs to the class of water-soluble B vitamins synthesized by the microbiotic flora in the digestive system of the animals (reticulorumen and large intestine). Owing to the microbes' own requirements, the synthesis usually does not produce sufficient quantities to cover the needs of the entire animal organism. Marked deficiencies occur rarely, even in case of an inadequate supply with cyanocobalamin."
TLS comment: We have heard of multiple sub-dermal B12 injections being given to horses during acupuncture treatments (may be called AIT - Acupuncture Injection Therapy or biopuncture). It is suggested that injecting a sterile solution containing e.g. vitamins at an acupuncture point may provide lingering stimulation. Whether this is true/effective or not, from a nutritional viewpoint humans with vitamin B12 deficiency may benefit from injections of vitamin B12 to boost their B12 levels, whereas horses will not as they have sufficient B12.
Vitamin B12 is a water-soluble vitamin and cannot be stored in the body (in significant amounts).
There is no dietary requirement for vitamin B12 above that supplied by intestinal synthesis.
Horses (only) need cobalt in their diet so that their gut bacteria can produce vitamin B12.
There appears to be enough cobalt in forage and typical horse diets to meet cobalt and therefore vitamin B12 requirements.
A healthy hindgut microbiome is important for vitamin B12 production. In cattle, a high forage diet was linked to 3 x more vitamin B12 production than a high starch diet.
Supplementation with B12 might be considered following antibiotics or NSAIDs, diet changes, GI illness e.g. diarrhoea, high-intensity training or stressful events, general ill health or reduced appetite, for young or very old horses - any time the hind gut microbiome might be adversely affected.
Neither deficiency nor toxicity of B12 has been described in horses.
Vitamin B12 is also called cobalamin. B12 is synthesised naturally by microorganisms during hind gut fermentation, and absorbed from the caecum and colon. Horses are thought to have no requirement for B12 supplementation. Plants do not contain B12. Synthesis of B12 requires cobalt.
Cobalt and B12 are important for:
energy metabolism, e.g. the formation of glucose from volatile fatty acids, protein synthesis and fat metabolism,
the synthesis of myelin which coats nerves,
the maturation of red blood cells in bone marrow.
Caple et al. 1982 examined several hundred horses and reported plasma vitamin B12 concentrations between 1.8 and 7.3 micrograms/L. Intramuscular injections of vitamin B12 were rapidly cleared from the plasma, and large amounts excreted in the faeces via the bile when vitamin B12 was administered intravenously to foals.
There is very little research about cobalt in horses. Systemic absorption was demonstrated from the hindgut after administration of cyanocobalamin cobalt into the caecum (Stillions 1971). Cobalt is absorbed as part of the cobalamin complex formed through bacterial synthesis, and a constant replenishment of cobalt is required for cobalamin synthesis. "Microbial synthesis of cobalamin by the equine intestinal community occurs and obviously appears to provide sufficient cobalamin for the horse (Salminen 1975)" (Source: Equine Applied and Clinical Nutrition 2013). 0.1 mg/kg DM diet is considered adequate (McDowell 2003). Sources of cobalt: grass pasture 0.5-0.2 mg/kg DM, alfalfa 0.3 mg/kg DM, yeast is also a good source of cobalt. Straw is low in cobalt.
Source: Equine Applied and Clinical Nutrition 2013, NRC Nutrient Requirements of Horses 2007.
Vitamin B12 - Do Horses Need It In Their Diet? Nerida Richards
Can Vitamin B12 Improve a Horse's Appetite? Clair Thunes July 2018 www.thehorse.com
The Story of B Vitamins in Horse Nutrition - KER June 2012
Hillyer LL, Ridd Z, Fenwick S, Hincks P, Paine SW.
Pharmacokinetics of inorganic cobalt and a vitamin B12 supplement in the Thoroughbred horse: Differentiating cobalt abuse from supplementation.
Equine Vet J. 2018 May;50(3):343-349. doi: 10.1111/evj.12774. Epub 2017 Nov 12. PMID: 29053883.
Wenzel R, Major D, Hesp K, Doble P.
Determination of vitamin B12 in equine urine by liquid chromatography - inductively coupled plasma - mass spectrometry.
J Trace Elem Med Biol. 2018 Dec;50:634-639. doi: 10.1016/j.jtemb.2018.05.005. Epub 2018 May 9. PMID: 29773468.
"a horse can excrete a significant proportion of a commercially available vitamin B12 injection in urine without metabolic modification."
Roberts MC.
Serum and red cell folate and serum vitamin B12 levels in horses.
Aust Vet J. 1983 Apr;60(4):106-11. doi: 10.1111/j.1751-0813.1983.tb05906.x.
Vitamin B12 injections
Vitbee is an injection for horses and cattle containing 0.100% w/v cyanocobalamin.
"Indications for use, specifying the target species: As an aid in raising Vitamin B12 levels in cattle and horses, calves and foals. For use in the treatment of Vitamin B12 deficiency, and for poor growth rates and general unthriftiness in young animals when associated with the above deficiency." "Following treatment, the vitamin B12 status of herbivores should be maintained by dietary supplementation with cobalt."
Butasal is an injection for horses, cattle, dogs and cats containing phosphorus as Butafosfan and 0.05 mg cyanocobalamin (vitamin B12) per ml.
"Cyanocobalamin is a co-enzyme in the biosynthesis of glucose from propionate. Further it serves as a co-factor to enzymes important in fatty acid synthesis and is important for maintenance of normal haemopoiesis, protection of the liver, and maintenance of muscle tissue, healthy skin, brain and pancreatic metabolism. It belongs to the class of water-soluble B vitamins synthesized by the microbiotic flora in the digestive system of the animals (reticulorumen and large intestine). Owing to the microbes' own requirements, the synthesis usually does not produce sufficient quantities to cover the needs of the entire animal organism. Marked deficiencies occur rarely, even in case of an inadequate supply with cyanocobalamin."
TLS comment: We have heard of multiple sub-dermal B12 injections being given to horses during acupuncture treatments (may be called AIT - Acupuncture Injection Therapy or biopuncture). It is suggested that injecting a sterile solution containing e.g. vitamins at an acupuncture point may provide lingering stimulation. Whether this is true/effective or not, from a nutritional viewpoint humans with vitamin B12 deficiency may benefit from injections of vitamin B12 to boost their B12 levels, whereas horses will not as they have sufficient B12.
Vitamin C
What is vitamin C and what is it needed for?
Vitamin C (L-ascorbic acid and dehydro-L-ascorbic acid) is a water soluble, non-enzymatic antioxidant and is involved in preventing radical attacks, regenerating other antioxidants e.g. vitamin E, hormone (norepinephrine) synthesis, collagen synthesis, carnitine synthesis, the hydroxylation of proline and lysine, bone calcification, protecting the airways and maintaining respiratory function, immune system, wound healing, clotting function, joint function, cartilage formation.
Decreased blood ascorbic acid levels have been seen in horses with significant wounds (including post-surgery), epistaxis (nose bleeds), strangles, flu, acute rhinopneumonia, recurrent airway obstruction, performance insufficiency, during transport, and age (being older than 20 years) and intensive exercise may affect blood levels. Vitamin C is decreased in the lung lining fluid of horses with airway inflammation.
Vitamin C increases iron absorption.
Blood levels of vitamin C
Ascorbic acid can be measured in the blood (plasma or serum). Levels recorded in research range from 0.8 to 6.5 ug/ml (or mg/l), and have been reported to be lower in winter than in summer:
5.9 ug/ml in 488 healthy adult horses - Jaeschke and Keller 1978
2 - 4.3 ug/ml in 20 unsupplemented Thoroughbred racehorses from February to October - Snow et al. 1987
0.8 - 4.6 ug/ml in endurance racing horses - Hargreaves et al. 2002, Marlin et al. 2002, Williams et al. 2004
3.2 ug/ml in unsupplemented Shetland ponies - Pearson et al. 1943
Where do horses get vitamin C from?
Horses appear to be able to synthesize vitamin C in the liver from glucose.
Several forms of vitamin C can be given to horses, including:
Ascorbyl palmitate - most efficient at raising blood ascorbic acid levels (i.e. most bioavailable)
Ascorbyl-2-monophosphate - slightly less effective than ascorbyl palmitate but slightly more effective than acetic acid at raising blood ascorbic acid levels, and may be the most stable form.
Ascorbic acid - least effective at raising blood ascorbic acid levels.
Snow et al. 1987 found that blood levels of vitamin C were significantly higher after supplementing with calcium ascorbyl-2-monophosphate and ascorbyl palmitate rather than ascorbic acid, and Snow and Frig 1989 found that ascorbyl palmitate had greater bioavailability than ascorbic acid.
Dr David Marlin of Science Supplements wrote: "With reference to Vitamin C, supplements that contain L-ascorbic acid are very poorly absorbed by horses so you should be looking for a supplement which includes Ascorbyl Monophosphate as the source of Vitamin C (Deaton, Marlin, Smith et al., 2003). Ascorbyl palmitate is also well absorbed by horses but is not particularly stable in supplements." He refers to Deaton et al. 2003 (see below) which found the following mean plasma ascorbic acid concentrations after 2 weeks of supplementation: ascorbyl palmitate 29, ascorbyl monophosphate 23, control 18 mmol/l (the paper does not explain whether control means no supplemental vitamin C at all, or 20 mg/kg bw ascorbic acid).
Spring Associated Respiratory Disease posted 31 March 2017.
So which form of vitamin C is best to feed? We are researching this...
Sources of vitamin C include:
Ascorbic acid:
PlusVital Vitamin C from Farm & Stable £21.74 per kg (Sept 2019). 100% ascorbic acid. 22 p per 10 g vitamin C.
Vit C Equine from Aviform £19.95 per kg (Sept 2019). 99.5% ascorbic acid. 20 p per 10 g vitamin C (however, this product was out of stock in Sept 2019 and Aviform could not say when it would be back in stock).
Ascorbyl monophosphate
Pure Vitamin C from Equestrizone £64.95 for 3 kg = £21.65 per kg (Sept 2019). 35% sodium calcium ascorbyl-2-phosphate (3a311). 67 p per 10 g vitamin C.
Always check the actual amount of the ingredient in a supplement, and if not 100%, make sure you know what makes up the difference. This product C-Vite from Animal Health Company contains 12.5% ascorbic acid and does not describe the other 87.5% of the product. At £15 per kg product (Sept 2019), 10 g vitamin C would cost £1.20!
Vitamin C requirements
A recommended daily amount has not been established for vitamin C. Healthy horses are assumed to meet their needs by internal synthesis, and there is currently no scientific basis for regularly supplementing healthy horses with vitamin C. Vitamin C deficiency has not been reported.
Horses suffering from the conditions listed above that have been shown to lower blood vitamin C levels may benefit from oral supplementation. Kolb et al. 1983 recommended 30 mg/kg bodyweight of vitamin C (ascorbyl-2-triphosphate) (15 g per day for a 500 kg horse). If supplemental vitamin C is given, it must not be stopped suddenly.
Too much vitamin C
There is little information about the effects of excess vitamin C in horses, and toxicity has not been reported. In other speciess excess vitamin C has caused allergic reactions and decreased natural synthesis of vitamin C. Excess amounts could act as a metabolic acidifier, pro-oxidant and cause gastro-intestinal disturbance. 44 mg ascorbic acid per kg bodyweight has been given to horses daily for 8 months with no recorded negative effects (Snow et al. 1987).
More information about vitamin C
Ingested Vitamin C enters the plasma and extracellular fluid relatively quickly, and is then taken up by various cell types, including white blood cells. It has been suggested that serum vitamin C levels will only increase when the vitamin C stores of other cells have become saturated, which may explain why an increase in blood vitamin C levels are not always seen at the start of supplementation.
Vit C and PPID - The Laminitis Site
Vitamin C – Friend or Foe - Dr Eleanor Kellon, May 2019
Vitamin C for Horses - Dr Eleanor Kellon Oct 2017
O'Callaghan DK, Schall SA, Birmingham SSW, Lehman JS.
Protective Effects of Ascorbic Acid and α-Tocopherol on the In Vitro Oxidation of Equine Erythrocytes Caused by Extracts of Wilted Red Maple Leaves.
Journal of Equine Veterinary Science Nov-Dec 2015 Volume 35, Issues 11–12 Pages 940-946, https://doi.org/10.1016/j.jevs.2015.08.022.
Horses supplemented daily for 12 weeks with ascorbic acid had reduced erythrocyte oxidation following red maple leaf toxicosis (compared to horses given no supplement, and less than horses given vitamin E (alpha-tocopherol).
Vitamin C for Horses - Kentucky Equine Research July 2015
Horses can synthesize vitamin C from glucose in the liver, and fresh forage e.g. grass will easily meet vitamin C requirements, but hay contains low levels of vitamin C due to oxidation, and commercial feeds tend to be fairly low in vitamin C as potency decreases after manufacture. Deficiency is very unusual, but excessive stress may reduce vitamin C synthesis and supplementation may be necessary. "Due to lack of intestinal absorption, more than three grams per day of vitamin C is necessary to elicit a change in blood ascorbate levels".
Yaqub LS, Mshelia WP, Ayo JO
Erythrocyte Osmotic Fragility and Hematological Responses of horses Administered Ascorbic Acid and Exposed to Road Transportation
JEVS Nov-Dec 2014 Vol 34 Issues 11-12 pages 1324-1328. https://doi.org/10.1016/j.jevs.2014.09.015
Horses given 200 mg/kg ascorbic acid had reduced erythrocyte fragility and improved haematological parameters following 6 hours road transportation compared to controls.
Vitamin C in horse diets - Kentucky Equine Research December 2013
Bad Science Awards for Equine Supplements! Dr David Marlin - L-Ascorbic Acid (vitamin C)
"Supplementation with Vitamin C increases plasma, airway and tissue concentrations of Vitamin C leading to greater antioxidant function and associated benefits." L-ascorbic acid in supplements is poorly absorbed by horses and very unstable; more easily absorbed forms of Vitamin C include ascorbyl palmitate, ascorbyl sulphate or ascorbyl monophosphate.
Winther K, Kharazmi A, Hansen ASV, Falk-Rønne J
The absorption of natural vitamin C in horses and anti-oxidative capacity: a randomised, controlled study on trotters during a three-month intervention period
Comparative Exercise Physiology, 2012; 8 (3/4): 195-201
16 horses aged 2 to 6 in training were given either 25 g (~125 mg vitamin C) or 50 g (~250 mg vitamin C) of whole rosehip powder (LitoVet). No control group was used, no cross-over, no comparison with artificial vitamin C. Blood was tested on days 0, 14, 28 and 84 for vitamin C and oxidative anions.
The horses given 25 g a day had no significant increase in serum vitamin C levels (D0 20, D14 18.8, D28 20.1, D84 23.5 umol/L) and a small reduction in release of oxidative anions (D0 4498, D14 4340, D84 3351.
The horses given 50 g a day had a significant increase in serum vitamin C levels (D0 18.5, D14 21.4, D28 23.4, D84 26.7 umol/L) and a reduction in release of oxidative anions that was greater at day 14 than day 84 (D0 4304, D14 2276, D84 3397).
The researchers suggest that other anti-oxidants in the rosehip (e.g. vitamin E, beta-carotene, GOPO) may have contributed to the reduction in the release of oxidative anions.
Conclusion: long-term supplementation of 250 mg of natural vitamin C from whole dried rosehips improves serum vitamin C levels comparable to those obtained using 80 x the amount of synthetic vitamin C as ascorbic acid (Loscher et al. 1984, Snow et al. 1987), perhaps due to co-factors within the rosehip that facilitate the uptake of vitamin C from the digestive tract.
Winther et al. 2010 found that a single administration of 1000 mg of natural vitamin C, when fed as 210 g of rosehip powder resulted in a significant increase in the serum concentration of vitamin C in horses after just 2 hours.
Winther K, Kharazmi A, Hansen ASV, Falk-Rønne J
A randomised placebo controlled double blind study on the effect of subspecies of rose hip (Rosa canina) on the immune system, working capacity and behaviour of horses
In: Ellis AD, Longland AC, Coenen M, Miraglia N, The impact of nutrition on the health AND welfare of horses, EAAP volume 128 2010. Wageningen Academic Publishers, Wageningen, the Neteherlands, pp. 283-287.
Deaton et al. 2004 and Kirschvink et al. 2002 reported the importance of pulmonary vitamin C status in horses with recurrent airway obstruction.
Deaton CM, Marlin DJ, Smith NC, Harris PA, Roberts CA, Schroter RC, Kelly FJ
Pulmonary epithelial lining fluid and plasma ascorbic acid concentrations in horses affected by recurrent airway obstruction
Am J Vet Res. 2004 Jan;65(1):80-7
Deaton et al. 2003 established that the vitamin C content of lung lining fluid is influenced by the vitamin C content of the diet.
Deaton CM, Marlin DJ, Smith NC, Roberts CA, Harris PA, Kelly FJ, Schroter RC
Pulmonary bioavailability of ascorbic acid in an ascorbate-synthesising species, the horse
Free Radic Res. 2003 Apr;37(4):461-7
6 healthy ponies were given 3 x 2 week treatments: AP = oral vitamin C as ascorbyl palmitate equivalent to 20 mg ascorbic acid/kg bw; CAM = oral vitamin C as calcium ascorbyl-2-monophosphate equivalent to 20 mg ascorbic acid/kg bw; C = control (no treatment).
After 2 weeks mean plasma ascorbic acid concentrations were AP 29, CAM 23, C 18 mmol/l.
After 2 weeks, the concentrations of ascorbic acid in bronchoalveolar lavage fluid were AP 30, CAM 25, C 18 mmol/l - increases were seen in only 5/6 ponies.
Conclusion: "the concentration of lung lining fluid ascorbic acid is increased following ascorbic acid supplementation (20 mg/kg body weight) in an ascorbate-synthesising species."
What is vitamin C and what is it needed for?
Vitamin C (L-ascorbic acid and dehydro-L-ascorbic acid) is a water soluble, non-enzymatic antioxidant and is involved in preventing radical attacks, regenerating other antioxidants e.g. vitamin E, hormone (norepinephrine) synthesis, collagen synthesis, carnitine synthesis, the hydroxylation of proline and lysine, bone calcification, protecting the airways and maintaining respiratory function, immune system, wound healing, clotting function, joint function, cartilage formation.
Decreased blood ascorbic acid levels have been seen in horses with significant wounds (including post-surgery), epistaxis (nose bleeds), strangles, flu, acute rhinopneumonia, recurrent airway obstruction, performance insufficiency, during transport, and age (being older than 20 years) and intensive exercise may affect blood levels. Vitamin C is decreased in the lung lining fluid of horses with airway inflammation.
Vitamin C increases iron absorption.
Blood levels of vitamin C
Ascorbic acid can be measured in the blood (plasma or serum). Levels recorded in research range from 0.8 to 6.5 ug/ml (or mg/l), and have been reported to be lower in winter than in summer:
5.9 ug/ml in 488 healthy adult horses - Jaeschke and Keller 1978
2 - 4.3 ug/ml in 20 unsupplemented Thoroughbred racehorses from February to October - Snow et al. 1987
0.8 - 4.6 ug/ml in endurance racing horses - Hargreaves et al. 2002, Marlin et al. 2002, Williams et al. 2004
3.2 ug/ml in unsupplemented Shetland ponies - Pearson et al. 1943
Where do horses get vitamin C from?
Horses appear to be able to synthesize vitamin C in the liver from glucose.
Several forms of vitamin C can be given to horses, including:
Ascorbyl palmitate - most efficient at raising blood ascorbic acid levels (i.e. most bioavailable)
Ascorbyl-2-monophosphate - slightly less effective than ascorbyl palmitate but slightly more effective than acetic acid at raising blood ascorbic acid levels, and may be the most stable form.
Ascorbic acid - least effective at raising blood ascorbic acid levels.
Snow et al. 1987 found that blood levels of vitamin C were significantly higher after supplementing with calcium ascorbyl-2-monophosphate and ascorbyl palmitate rather than ascorbic acid, and Snow and Frig 1989 found that ascorbyl palmitate had greater bioavailability than ascorbic acid.
Dr David Marlin of Science Supplements wrote: "With reference to Vitamin C, supplements that contain L-ascorbic acid are very poorly absorbed by horses so you should be looking for a supplement which includes Ascorbyl Monophosphate as the source of Vitamin C (Deaton, Marlin, Smith et al., 2003). Ascorbyl palmitate is also well absorbed by horses but is not particularly stable in supplements." He refers to Deaton et al. 2003 (see below) which found the following mean plasma ascorbic acid concentrations after 2 weeks of supplementation: ascorbyl palmitate 29, ascorbyl monophosphate 23, control 18 mmol/l (the paper does not explain whether control means no supplemental vitamin C at all, or 20 mg/kg bw ascorbic acid).
Spring Associated Respiratory Disease posted 31 March 2017.
So which form of vitamin C is best to feed? We are researching this...
Sources of vitamin C include:
Ascorbic acid:
PlusVital Vitamin C from Farm & Stable £21.74 per kg (Sept 2019). 100% ascorbic acid. 22 p per 10 g vitamin C.
Vit C Equine from Aviform £19.95 per kg (Sept 2019). 99.5% ascorbic acid. 20 p per 10 g vitamin C (however, this product was out of stock in Sept 2019 and Aviform could not say when it would be back in stock).
Ascorbyl monophosphate
Pure Vitamin C from Equestrizone £64.95 for 3 kg = £21.65 per kg (Sept 2019). 35% sodium calcium ascorbyl-2-phosphate (3a311). 67 p per 10 g vitamin C.
Always check the actual amount of the ingredient in a supplement, and if not 100%, make sure you know what makes up the difference. This product C-Vite from Animal Health Company contains 12.5% ascorbic acid and does not describe the other 87.5% of the product. At £15 per kg product (Sept 2019), 10 g vitamin C would cost £1.20!
Vitamin C requirements
A recommended daily amount has not been established for vitamin C. Healthy horses are assumed to meet their needs by internal synthesis, and there is currently no scientific basis for regularly supplementing healthy horses with vitamin C. Vitamin C deficiency has not been reported.
Horses suffering from the conditions listed above that have been shown to lower blood vitamin C levels may benefit from oral supplementation. Kolb et al. 1983 recommended 30 mg/kg bodyweight of vitamin C (ascorbyl-2-triphosphate) (15 g per day for a 500 kg horse). If supplemental vitamin C is given, it must not be stopped suddenly.
Too much vitamin C
There is little information about the effects of excess vitamin C in horses, and toxicity has not been reported. In other speciess excess vitamin C has caused allergic reactions and decreased natural synthesis of vitamin C. Excess amounts could act as a metabolic acidifier, pro-oxidant and cause gastro-intestinal disturbance. 44 mg ascorbic acid per kg bodyweight has been given to horses daily for 8 months with no recorded negative effects (Snow et al. 1987).
More information about vitamin C
Ingested Vitamin C enters the plasma and extracellular fluid relatively quickly, and is then taken up by various cell types, including white blood cells. It has been suggested that serum vitamin C levels will only increase when the vitamin C stores of other cells have become saturated, which may explain why an increase in blood vitamin C levels are not always seen at the start of supplementation.
Vit C and PPID - The Laminitis Site
Vitamin C – Friend or Foe - Dr Eleanor Kellon, May 2019
Vitamin C for Horses - Dr Eleanor Kellon Oct 2017
O'Callaghan DK, Schall SA, Birmingham SSW, Lehman JS.
Protective Effects of Ascorbic Acid and α-Tocopherol on the In Vitro Oxidation of Equine Erythrocytes Caused by Extracts of Wilted Red Maple Leaves.
Journal of Equine Veterinary Science Nov-Dec 2015 Volume 35, Issues 11–12 Pages 940-946, https://doi.org/10.1016/j.jevs.2015.08.022.
Horses supplemented daily for 12 weeks with ascorbic acid had reduced erythrocyte oxidation following red maple leaf toxicosis (compared to horses given no supplement, and less than horses given vitamin E (alpha-tocopherol).
Vitamin C for Horses - Kentucky Equine Research July 2015
Horses can synthesize vitamin C from glucose in the liver, and fresh forage e.g. grass will easily meet vitamin C requirements, but hay contains low levels of vitamin C due to oxidation, and commercial feeds tend to be fairly low in vitamin C as potency decreases after manufacture. Deficiency is very unusual, but excessive stress may reduce vitamin C synthesis and supplementation may be necessary. "Due to lack of intestinal absorption, more than three grams per day of vitamin C is necessary to elicit a change in blood ascorbate levels".
Yaqub LS, Mshelia WP, Ayo JO
Erythrocyte Osmotic Fragility and Hematological Responses of horses Administered Ascorbic Acid and Exposed to Road Transportation
JEVS Nov-Dec 2014 Vol 34 Issues 11-12 pages 1324-1328. https://doi.org/10.1016/j.jevs.2014.09.015
Horses given 200 mg/kg ascorbic acid had reduced erythrocyte fragility and improved haematological parameters following 6 hours road transportation compared to controls.
Vitamin C in horse diets - Kentucky Equine Research December 2013
Bad Science Awards for Equine Supplements! Dr David Marlin - L-Ascorbic Acid (vitamin C)
"Supplementation with Vitamin C increases plasma, airway and tissue concentrations of Vitamin C leading to greater antioxidant function and associated benefits." L-ascorbic acid in supplements is poorly absorbed by horses and very unstable; more easily absorbed forms of Vitamin C include ascorbyl palmitate, ascorbyl sulphate or ascorbyl monophosphate.
Winther K, Kharazmi A, Hansen ASV, Falk-Rønne J
The absorption of natural vitamin C in horses and anti-oxidative capacity: a randomised, controlled study on trotters during a three-month intervention period
Comparative Exercise Physiology, 2012; 8 (3/4): 195-201
16 horses aged 2 to 6 in training were given either 25 g (~125 mg vitamin C) or 50 g (~250 mg vitamin C) of whole rosehip powder (LitoVet). No control group was used, no cross-over, no comparison with artificial vitamin C. Blood was tested on days 0, 14, 28 and 84 for vitamin C and oxidative anions.
The horses given 25 g a day had no significant increase in serum vitamin C levels (D0 20, D14 18.8, D28 20.1, D84 23.5 umol/L) and a small reduction in release of oxidative anions (D0 4498, D14 4340, D84 3351.
The horses given 50 g a day had a significant increase in serum vitamin C levels (D0 18.5, D14 21.4, D28 23.4, D84 26.7 umol/L) and a reduction in release of oxidative anions that was greater at day 14 than day 84 (D0 4304, D14 2276, D84 3397).
The researchers suggest that other anti-oxidants in the rosehip (e.g. vitamin E, beta-carotene, GOPO) may have contributed to the reduction in the release of oxidative anions.
Conclusion: long-term supplementation of 250 mg of natural vitamin C from whole dried rosehips improves serum vitamin C levels comparable to those obtained using 80 x the amount of synthetic vitamin C as ascorbic acid (Loscher et al. 1984, Snow et al. 1987), perhaps due to co-factors within the rosehip that facilitate the uptake of vitamin C from the digestive tract.
Winther et al. 2010 found that a single administration of 1000 mg of natural vitamin C, when fed as 210 g of rosehip powder resulted in a significant increase in the serum concentration of vitamin C in horses after just 2 hours.
Winther K, Kharazmi A, Hansen ASV, Falk-Rønne J
A randomised placebo controlled double blind study on the effect of subspecies of rose hip (Rosa canina) on the immune system, working capacity and behaviour of horses
In: Ellis AD, Longland AC, Coenen M, Miraglia N, The impact of nutrition on the health AND welfare of horses, EAAP volume 128 2010. Wageningen Academic Publishers, Wageningen, the Neteherlands, pp. 283-287.
Deaton et al. 2004 and Kirschvink et al. 2002 reported the importance of pulmonary vitamin C status in horses with recurrent airway obstruction.
Deaton CM, Marlin DJ, Smith NC, Harris PA, Roberts CA, Schroter RC, Kelly FJ
Pulmonary epithelial lining fluid and plasma ascorbic acid concentrations in horses affected by recurrent airway obstruction
Am J Vet Res. 2004 Jan;65(1):80-7
Deaton et al. 2003 established that the vitamin C content of lung lining fluid is influenced by the vitamin C content of the diet.
Deaton CM, Marlin DJ, Smith NC, Roberts CA, Harris PA, Kelly FJ, Schroter RC
Pulmonary bioavailability of ascorbic acid in an ascorbate-synthesising species, the horse
Free Radic Res. 2003 Apr;37(4):461-7
6 healthy ponies were given 3 x 2 week treatments: AP = oral vitamin C as ascorbyl palmitate equivalent to 20 mg ascorbic acid/kg bw; CAM = oral vitamin C as calcium ascorbyl-2-monophosphate equivalent to 20 mg ascorbic acid/kg bw; C = control (no treatment).
After 2 weeks mean plasma ascorbic acid concentrations were AP 29, CAM 23, C 18 mmol/l.
After 2 weeks, the concentrations of ascorbic acid in bronchoalveolar lavage fluid were AP 30, CAM 25, C 18 mmol/l - increases were seen in only 5/6 ponies.
Conclusion: "the concentration of lung lining fluid ascorbic acid is increased following ascorbic acid supplementation (20 mg/kg body weight) in an ascorbate-synthesising species."
Vitamin D
Vitamin D has 2 forms:
D2 - ergocalciferol - 25-hydroxyvitamin D2 (25OHD2) - comes from plants exposed to sunlight;
D3 - cholecalciferol - 25-hydroxyvitamin D3 (25OHD3) - comes from skin exposure to sunlight.
Both forms are thought to be absorbed at a similar rate (around 60 to 80% absorption) but this figure is not from horse specific research (it's from Schenk & Kolb 1982). McDowell 2000 found that vitamin D levels in compound feed deteriorated by 10 to 30 % when stored for 4 or 6 months at 22'C.
What is known about vitamin D in other mammals may not always relate to horses. For example, in most mammals, vitamin D is important for calcium and phosphorus regulation, but in horses the importance of vitamin D is questionable. And recent research from New Zealand has suggested that horses may need to get vitamin D from their diet, rather than exposure to sunlight. So when reading about vitamin D, make sure any research cited is specific to horses.
Sun-dried hay and haylage will contain higher levels of vitamin D than barn/artificially dried forage - Ballet et al. 2000 found around 971 IU/kg DM vitamin D in sun dried hay v 470 IU/kg DM in barn dried hay. Leaves store the most vitamin D, so forage with a higher leaf to stem ratio will contain more vitamin D. Ballet et al. 2000 gave ranges for vitamin D in forage:
fresh green forage 31 to 1800 IU/kg DM
dried lucerne 176 to 617 IU/kg DM
hay 90 to 5560 IU/kg DM.
The NRC requirement for vitamin D is 3300 IU for all classes of adult 500 kg horse.
Do Horses Need Vitamin D Supplemenation? Clair Thunes, October 2020, www.thehorse.com
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
Dosi MCMC, McGorum BC, Kirton RD, Cillan-Garcia E, Mellanby RJ, Keen JA, Hurst EA, Morgan RA
The effect of season, management and endocrinopathies on vitamin D status in horses
Equine Veterinary Journal August 2022 DOI: 10.1111/evj.13873
Horse vitamin D (25(OH)D) concentrations are exceptionally low compared with other species, and considerably below the threshold indicative of deficiency in humans (which would be associated with a high risk for the development of nutritional osteomalacia). Nutritional osteomalacia has not been reported in horses, and developmental rickets has only been reported anecdotally or induced in experimental settings. It is suggested that calcium metabolism in horses is influenced by a lesser extent by vitamin D status than in other species. Endogenous cutaneous production of vitmin D3 in horses is either non-existent or minimal, vitamin D concentration is mostly influenced by eating vitamin D2 in plants or supplementation with D2 or D3. Exercise has been shown to reduce plasma 25(OH)D2 levels. Vitamin D is stored inside adipose tissue. Obese horses were found to have higher 25(OH)D2 levels; the authors speculated that adipose tissue inflammation and dysfunction in obese horses could impair uptake and use of vitamin D from fat stores, leading to higher circulating vitamin D2 levels [TLS: or could it simply be that obese horses ate more forage, and hence more vitamin D2!]. No association was found between insulin concentrations and vitamin D status. There appears to be no direction relationship between ACTH (PPID) and vitamin D levels.
This research suggests that grass pasture/forage is likely to meet horse vitamin D requirements, and supplementation of vitamin D is unlikely to be necessary for horses with access to pasture/forage. However more research is needed to establish requirements for hard working horses with limited access to pasture or forage.
See also Vitamin D Biology Cannot Be Extrapolated from Humans to horses - KER November 2022
Azarpeykan S, Dittmer KE, Gee EK, Marshall JC, Wallace J, Elder P, Acke E, Thompson KG
Influence of blanketing and season on vitamin D and parathyroid hormone, calcium, phosphorus, and magnesium concentrations in horses in New Zealand
Domest Anim Endocrinol. 2016 Jul;56:75-84. doi: 10.1016/j.domaniend.2016.03.003. Epub 2016 Mar 19
Of 21 healthy horses kept at pasture for 13 months, 5 were covered with standard rugs with neck covers, and 16 horses left uncovered. "The serum concentration of 25OHD3 in horses was either undetectable or below the detection limit of the assay, and the main form of 25OHD was 25OHD2. No differences in serum 25OHD2, 1,25(OH)2D, iCa, tCa, P, tMg, and PTH (P ≥ 0.05) concentrations were seen between the 2 groups. Associations were seen between iCa and PTH (P < 0.05), iCa and tMg (P < 0.05), and dietary vitamin D and 25OHD2 (P < 0.05). A strong seasonal trend was seen in serum 25OHD2 (P < 0.0001), which was higher during spring and summer when the amount of sunshine and UV radiation was higher. Parathyroid hormone and 1,25(OH)2D showed opposing trends with PTH higher in winter whereas 1,25(OH)2D was higher in summer. The results suggest that dietary vitamin D may be necessary for horses to fulfill their vitamin D requirements; however, further research is required to determine the contribution of vitamin D3 synthesis in the skin to the vitamin D status of the horse."
Azarpeykan, S., Gee, EK., Thompson, KG., Acke, E., Marshall, JC., & Dittmer, KE. (2014).
Vitamin d status of covered and non-covered horses at pasture.
Proceedings of the Australasian Equine Science Symposium. Vol. 5 (pp. 33).
"Many factors are recognized as having a major influence on the ability of skin to synthesise vitamin D3 including dermal melanin pigmentation, season, latitude and altitude, type of clothing, aging and the timing of sunlight exposure. The importance of skin covering on vitamin D3 synthesis is well documented in human literature. There is a deficit in the equine literature regarding vitamin D synthesis and factors affecting its metabolism. The majority of horses in New Zealand have dark skin and spend a large proportion of their time outside in paddocks often with covers on, both of which may adversely affect their ability to synthesise vitamin D3 in the skin. The purpose of the present study was to determine whether horses that are covered for substantial periods of time have lower serum vitamin D concentrations than horses that are not covered. Twenty one mature horses were included in the study. Five were covered with standard horse rugs, including a neck rug. All horses were fed ad libitum on grass pasture, and hay when appropriate. Blood and pasture samples were collected monthly (January-June 2013) and analysed for total 25- hydroxyvitamin D and 1,25-dihydroxyvitamin D."
Dittmer, K. (2015). Do Blanketed Horses Get Enough Vitamin D?
"Many horses today have wardrobes that rival those of their owners-winter rugs, turnout sheets, rain covers, fly gear, and more. We know these blankets can help keep horses warm, dry, and fly-free, but how they impact some aspects of horse health remain unclear. Take vitamin D, for example: Horses need sunlight to synthesize this vitamin that's important for bone health. So does blanketing affect their ability to produce it? That's what a research team from New Zealand recently tried to find out."
Do Blanketed Horses Get Enough Vitamin D? - Erica Larson, www.thehorse.com June 2015
El Shorafa WM, Feaster JP, Ott EA, Asquith RL.
Effect of vitamin D and sunlight on growth and bone development of young ponies.
J Anim Sci. 1979 Apr;48(4):882-6. doi: 10.2527/jas1979.484882x. PMID: 479023.
Vitamin D has 2 forms:
D2 - ergocalciferol - 25-hydroxyvitamin D2 (25OHD2) - comes from plants exposed to sunlight;
D3 - cholecalciferol - 25-hydroxyvitamin D3 (25OHD3) - comes from skin exposure to sunlight.
Both forms are thought to be absorbed at a similar rate (around 60 to 80% absorption) but this figure is not from horse specific research (it's from Schenk & Kolb 1982). McDowell 2000 found that vitamin D levels in compound feed deteriorated by 10 to 30 % when stored for 4 or 6 months at 22'C.
What is known about vitamin D in other mammals may not always relate to horses. For example, in most mammals, vitamin D is important for calcium and phosphorus regulation, but in horses the importance of vitamin D is questionable. And recent research from New Zealand has suggested that horses may need to get vitamin D from their diet, rather than exposure to sunlight. So when reading about vitamin D, make sure any research cited is specific to horses.
Sun-dried hay and haylage will contain higher levels of vitamin D than barn/artificially dried forage - Ballet et al. 2000 found around 971 IU/kg DM vitamin D in sun dried hay v 470 IU/kg DM in barn dried hay. Leaves store the most vitamin D, so forage with a higher leaf to stem ratio will contain more vitamin D. Ballet et al. 2000 gave ranges for vitamin D in forage:
fresh green forage 31 to 1800 IU/kg DM
dried lucerne 176 to 617 IU/kg DM
hay 90 to 5560 IU/kg DM.
The NRC requirement for vitamin D is 3300 IU for all classes of adult 500 kg horse.
Do Horses Need Vitamin D Supplemenation? Clair Thunes, October 2020, www.thehorse.com
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
Dosi MCMC, McGorum BC, Kirton RD, Cillan-Garcia E, Mellanby RJ, Keen JA, Hurst EA, Morgan RA
The effect of season, management and endocrinopathies on vitamin D status in horses
Equine Veterinary Journal August 2022 DOI: 10.1111/evj.13873
Horse vitamin D (25(OH)D) concentrations are exceptionally low compared with other species, and considerably below the threshold indicative of deficiency in humans (which would be associated with a high risk for the development of nutritional osteomalacia). Nutritional osteomalacia has not been reported in horses, and developmental rickets has only been reported anecdotally or induced in experimental settings. It is suggested that calcium metabolism in horses is influenced by a lesser extent by vitamin D status than in other species. Endogenous cutaneous production of vitmin D3 in horses is either non-existent or minimal, vitamin D concentration is mostly influenced by eating vitamin D2 in plants or supplementation with D2 or D3. Exercise has been shown to reduce plasma 25(OH)D2 levels. Vitamin D is stored inside adipose tissue. Obese horses were found to have higher 25(OH)D2 levels; the authors speculated that adipose tissue inflammation and dysfunction in obese horses could impair uptake and use of vitamin D from fat stores, leading to higher circulating vitamin D2 levels [TLS: or could it simply be that obese horses ate more forage, and hence more vitamin D2!]. No association was found between insulin concentrations and vitamin D status. There appears to be no direction relationship between ACTH (PPID) and vitamin D levels.
This research suggests that grass pasture/forage is likely to meet horse vitamin D requirements, and supplementation of vitamin D is unlikely to be necessary for horses with access to pasture/forage. However more research is needed to establish requirements for hard working horses with limited access to pasture or forage.
See also Vitamin D Biology Cannot Be Extrapolated from Humans to horses - KER November 2022
Azarpeykan S, Dittmer KE, Gee EK, Marshall JC, Wallace J, Elder P, Acke E, Thompson KG
Influence of blanketing and season on vitamin D and parathyroid hormone, calcium, phosphorus, and magnesium concentrations in horses in New Zealand
Domest Anim Endocrinol. 2016 Jul;56:75-84. doi: 10.1016/j.domaniend.2016.03.003. Epub 2016 Mar 19
Of 21 healthy horses kept at pasture for 13 months, 5 were covered with standard rugs with neck covers, and 16 horses left uncovered. "The serum concentration of 25OHD3 in horses was either undetectable or below the detection limit of the assay, and the main form of 25OHD was 25OHD2. No differences in serum 25OHD2, 1,25(OH)2D, iCa, tCa, P, tMg, and PTH (P ≥ 0.05) concentrations were seen between the 2 groups. Associations were seen between iCa and PTH (P < 0.05), iCa and tMg (P < 0.05), and dietary vitamin D and 25OHD2 (P < 0.05). A strong seasonal trend was seen in serum 25OHD2 (P < 0.0001), which was higher during spring and summer when the amount of sunshine and UV radiation was higher. Parathyroid hormone and 1,25(OH)2D showed opposing trends with PTH higher in winter whereas 1,25(OH)2D was higher in summer. The results suggest that dietary vitamin D may be necessary for horses to fulfill their vitamin D requirements; however, further research is required to determine the contribution of vitamin D3 synthesis in the skin to the vitamin D status of the horse."
Azarpeykan, S., Gee, EK., Thompson, KG., Acke, E., Marshall, JC., & Dittmer, KE. (2014).
Vitamin d status of covered and non-covered horses at pasture.
Proceedings of the Australasian Equine Science Symposium. Vol. 5 (pp. 33).
"Many factors are recognized as having a major influence on the ability of skin to synthesise vitamin D3 including dermal melanin pigmentation, season, latitude and altitude, type of clothing, aging and the timing of sunlight exposure. The importance of skin covering on vitamin D3 synthesis is well documented in human literature. There is a deficit in the equine literature regarding vitamin D synthesis and factors affecting its metabolism. The majority of horses in New Zealand have dark skin and spend a large proportion of their time outside in paddocks often with covers on, both of which may adversely affect their ability to synthesise vitamin D3 in the skin. The purpose of the present study was to determine whether horses that are covered for substantial periods of time have lower serum vitamin D concentrations than horses that are not covered. Twenty one mature horses were included in the study. Five were covered with standard horse rugs, including a neck rug. All horses were fed ad libitum on grass pasture, and hay when appropriate. Blood and pasture samples were collected monthly (January-June 2013) and analysed for total 25- hydroxyvitamin D and 1,25-dihydroxyvitamin D."
Dittmer, K. (2015). Do Blanketed Horses Get Enough Vitamin D?
"Many horses today have wardrobes that rival those of their owners-winter rugs, turnout sheets, rain covers, fly gear, and more. We know these blankets can help keep horses warm, dry, and fly-free, but how they impact some aspects of horse health remain unclear. Take vitamin D, for example: Horses need sunlight to synthesize this vitamin that's important for bone health. So does blanketing affect their ability to produce it? That's what a research team from New Zealand recently tried to find out."
Do Blanketed Horses Get Enough Vitamin D? - Erica Larson, www.thehorse.com June 2015
El Shorafa WM, Feaster JP, Ott EA, Asquith RL.
Effect of vitamin D and sunlight on growth and bone development of young ponies.
J Anim Sci. 1979 Apr;48(4):882-6. doi: 10.2527/jas1979.484882x. PMID: 479023.
Vitamin E
What is vitamin E - what does it do?
Fat soluble vitamin E (α-tocopherol) is essential for optimum health.
Vitamin E is an antioxidant - it protects cells from oxidative damage and traps reactive oxygen species to prevent free radical induced damage - and it is also important for normal neuromuscular, immune, cardiovascular, circulatory and reproductive function.
Vitamin E deficiency in young horses can cause equine neuroaxonal dystrophy (eNAD), equine degenerative myeloencephalopathy (EDM), and nutritional myodegeneration if selenium is also deficient.
Vitamin E deficiency in adult horses can cause equine motor neuron disease (EMND) and vitamin E deficient myopathy.
Vitamin E requirements - which horses need to have vitamin E supplemented?
The NRC Nutrient Requirements of Horses recommends 1 (maintenance) to 2 (hard work) IU vitamin E per kg bodyweight (500 - 1000 mg per 500 kg horse), with an upper safe limit of 20 IU per kg bodyweight (10,000 IU per 500 kg horse) synthetic vitamin E, above which coagulopathy, impaired bone mineralization and interference with absorption of other nutrients have been reported. Some nutritionists (including Pat Harris) recommend supplementing 2 mg/kg bodyweight vitamin E to all horses but this is not currently supported by research (source: Equine Applied and Clinical Nutrition 2013). Horses in intense training may benefit from 4 mg/kg bodyweight per day, and research has suggested that 1.5 to 5 mg/kg bodyweight vitamin E per day may be required to maintain vitamin E status in exercising horses. Schubert and Heinrich 2988 found superior race performance in Thoroughbreds supplementing with 1000 mg vitamin E per day on top of a basic diet that provided around 0.2 mg/kg bodyweight vitamin E.
Green grass contains plenty of vitamin E and horses with good access to fresh grass are unlikely to need vitamin E supplementation, but horses with no or restricted access to grass or access to brown grass (e.g. during drought) are likely to benefit from vitamin E supplementation, as vitamin E levels in hay and preserved forages decrease with time.
Horses eating high fat diets should have increased amounts of vitamin E added to their diet - 100-150 mg vitamin E is suggested for every 100 ml of oil added to the diet, or supplement 2 mg/kg bodyweight.
Exercise increases production of free radicals, so levels of antioxidants including vitamin E should increase accordingly to prevent signs of oxidative damage in working horses such as sore muscles, stiffness, and slow recovery from hard work.
Natural v synthetic
In humans, according to the National Institutes of Health "One mg vitamin E (alpha-tocopherol) (natural) is equivalent to 1 mg RRR-alpha-tocopherol (natural) or 2 mg all rac-alpha-tocopherol (synthetic)."
IU and mg
From 2019 the form of vitamin E should be listed in ingredients/analyses (not just "vitamin E") as either mg or mgTE or alpha-TE (alpha-tocopherol equivalents), a term established to account for the differences in biological activity of the various forms of vitamin E.
1 mg of alpha-tocopherol = 1 TE or 1 mgTE.
1 mg of natural RRR-alpha-Tocopherol = 1 mgTE.
1 mg of synthetic All-rac-alpha-Tocopherol = 0.5 mgTE.
1 IU of natural vitamin E including ester forms (d- or RRR-alpha tocopherol, RRR-alpha-tocopheryl acetate, RRR-alpha-tocopheryl succinate) = 0.67 mg. 1 mg natural vitamin E = 1.49 IU.
1 IU of synthetic vitamin E including ester forms (dl- or all-rac-alpha tocopherol, all rac-alpha-tocopheryl acetate, all rac-alpha-tocopheryl succinate) = 0.45 mg. 1 mg synthetic vitamin E = 2.22 IU.
(Source: Nestle Quality Assurance Centre Vitamin A, E & D Unit Change)
Typical amounts of vitamin E in the diet
Fresh forage and immature forage hay/haylage can contain 30-100 IU/kg DM vitamin E. Vitamin E levels in hay and haylage can also be very low. Cereals typically contain 20-30 IU/kg DM.
Vitamin E is destroyed by oxidation, and most food processes reduce vitamin E - milling, flaking, shredding, harvesting, drying and storing hay. High temperatures and humidity exacerbate vitamin E loss.
Absorption - which form of vitamin E should be used?
Vitamin E is supplemented as alpha-tocopherol. RRR-alpha tocopheral (also called d-alpha tocopheral) is natural vitamin E. all-rec-alpha tocopheral (also called dl-alpha tocopherol) is synthetic vitamin E. Research has suggested that bioavailability (how easily the vitamin E is absorbed from the digestive tract) is lowest with synthetic forms, better with natural acetate and alcohol forms, and best with water-soluble forms like Nano-e, which has been shown to rapidly increase blood levels of vitamin E. Natural d-alpha tocopherol is unstable, and therefore esterified, i.e. joined to an acid like acetate, to protect the anti-oxidant properties of the alpha-tocopherol, and released for absorption by enzymes during digestion.
Natural vitamin E RRR-alpha tocopherol has the highest biological activity at 1.49 IU per mg, with synthetic all-rac-alpha tocopherol having 1.10 IU/mg. Supplemental vitamin E is in ester form, with RRR-alpha-tocopheral acetate having 1.36 IU per mg and all-rac-alpha tocopheral acetate 1.00 IU per mg (source: NRC Nutrient Requirements of Horses 2007 and Equine Applied and Clinical Nutrition 2013).
Vitamin E is soluble in fat and absorption may be increased by feeding vitamin E mixed with oil.
Vitamin E is influenced by selenium status, and both vitamin E and selenium should be fed in appropriate amounts.
Fagan MM, Harris P, Adams A, Pazdro R, Krotky A, Call J, Duberstein KJ.
Form of Vitamin E Supplementation Affects Oxidative and Inflammatory Response in Exercising Horses.
J Equine Vet Sci. 2020 Aug;91:103103. doi: 10.1016/j.jevs.2020.103103. Epub 2020 Apr 29. PMID: 32684249.
How are body concentrations of vitamin E tested?
As absorption and utilization of supplemental vitamin E may be individual to each horse, it is sometimes recommended to ensure that the diet provides the NRC recommended amount of vitamin E, and then test blood vitamin E concentrations to establish whether further vitamin E supplementation is required.
Vitamin E (alpha-tocopherol)status can be tested in blood serum or plasma. Normal reference range concentrations of alpha-tocopherol in horse plasma and serum are:
>2 μg/mL adequate
1.5 - 2 μg/mL marginal
<2 μg/mL deficient
However, alpha-tocopherol concentrations can fluctuate significantly, and a single serum sample may be an unsatisfactory indicator of vitamin E status.
Vitamin E in Equine Nutrition: Three Questions - Stephanie Valberg, KER September 2019
Adult horse requirement is 1-2 IU vitamin E per kg bodyweight per day. Grazing fresh grass provided 350-3000 IU of vitamin E per day to horses eating 1.7% of their bodyweight of pasture grass, depending on the season. Vitamin E degrades with feed/forage storage. Supplemented vitamin E is either natural: RRR-alpha-tocopheral or synthetic: all-rac-alpha-tocopheral or dl-alpha-tocopherol. Natural has been shown to have higher activity than synthetic.
When Do Horses Need Vitamin E? Clair Thunes, August 2019, www.thehorse.com
Vitamin E in horses - UC Davis Veterinary Medicine
Vitamin E - Oregon State University (relating to humans)
Vitamin E – An Essential Nutrient for Horses - E Kane & RL Stuart
(Author bias: note Stuart Products sell natural vitamin E)
Vitamin E and Selenium Are Powerful Protectors - Dr Eleanor Kellon July 2018
Brown JC, Valberg SJ, Hogg M, Finno CJ
Effects of feeding two RRR-α-tocopherol formulations on serum, cerebrospinal fluid and muscle α-tocopherol concentrations in horses with subclinical vitamin E deficiency
Equine Vet J. 2017;49(6):753-758. doi:10.1111/evj.12692
Duberstein KJ, Pazdro R, Lee KC, Abrams A, Kane E, Stuart RL
Effect of Supplemental Vitamin E Form on Serum α-Tocopherol Levels and Blood Oxidative Stress Parameters in Response to a Novel Exercise Challenge
JEVS October 2017 Vol 57, Pages 61–66
(Author bias: note RL Stuart of Stuart Products sells natural vitamin E).
Finno CJ, Valberg SJ.
A comparative review of vitamin E and associated equine disorders.
J Vet Intern Med. 2012 Nov-Dec;26(6):1251-66. doi: 10.1111/j.1939-1676.2012.00994.x. Epub 2012 Aug 27. PMID: 22925200.
Pusterla N, Puschner B, Steidl S, Collier J, Kane E, Stuart RL
α-Tocopherol concentrations in equine serum and cerebrospinal fluid after vitamin E supplementation
Vet Rec. 2010;166:366–368
Synthetic all-rac-α-TP acetate given at 10,000 IU/day for 14 days increased serum α-TP from deficient to normal ranges, but had no impact on cerebrospinal fluid (CSF) α-TP concentrations.
Pagan JD, Kane E, Nash D
Form and source of tocopherol affects Vitamin E status in Thoroughbred horses
Pferdeheilkunde. 2005;21:101–102
Am J Vet Res. 1992 Dec;53(12):2228-34
Variability of alpha-tocopherol values associated with procurement, storage, and freezing of equine serum and plasma samples
Craig AM, Blythe LL, Rowe KE, Lassen ED, Barrington R, Walker KC
Alpha tocopherol levels in blood samples can be reduced by haemolysis (33% decrease), leaving the blood sample in contact with the rubber stopper of the collection tube during refrigeration for 72 hours (10% decrease), and storing blood at 20-25 degrees C for 72 hours (5% decrease).
Vitamin E in horses - UC Davis Veterinary Medicine
Bampidis et al.
Assessment of a feed additive consisting of all-rac-alpha-tocopheryl acetate (vitamin E) for all animal species for the renewal of its authorisation (NHU Europe GmbH)
EFSA Journal April 2021 Vol 19, Issue 4 https://doi.org/10.2903/j.efsa.2021.6533
What is vitamin E - what does it do?
Fat soluble vitamin E (α-tocopherol) is essential for optimum health.
Vitamin E is an antioxidant - it protects cells from oxidative damage and traps reactive oxygen species to prevent free radical induced damage - and it is also important for normal neuromuscular, immune, cardiovascular, circulatory and reproductive function.
Vitamin E deficiency in young horses can cause equine neuroaxonal dystrophy (eNAD), equine degenerative myeloencephalopathy (EDM), and nutritional myodegeneration if selenium is also deficient.
Vitamin E deficiency in adult horses can cause equine motor neuron disease (EMND) and vitamin E deficient myopathy.
Vitamin E requirements - which horses need to have vitamin E supplemented?
The NRC Nutrient Requirements of Horses recommends 1 (maintenance) to 2 (hard work) IU vitamin E per kg bodyweight (500 - 1000 mg per 500 kg horse), with an upper safe limit of 20 IU per kg bodyweight (10,000 IU per 500 kg horse) synthetic vitamin E, above which coagulopathy, impaired bone mineralization and interference with absorption of other nutrients have been reported. Some nutritionists (including Pat Harris) recommend supplementing 2 mg/kg bodyweight vitamin E to all horses but this is not currently supported by research (source: Equine Applied and Clinical Nutrition 2013). Horses in intense training may benefit from 4 mg/kg bodyweight per day, and research has suggested that 1.5 to 5 mg/kg bodyweight vitamin E per day may be required to maintain vitamin E status in exercising horses. Schubert and Heinrich 2988 found superior race performance in Thoroughbreds supplementing with 1000 mg vitamin E per day on top of a basic diet that provided around 0.2 mg/kg bodyweight vitamin E.
Green grass contains plenty of vitamin E and horses with good access to fresh grass are unlikely to need vitamin E supplementation, but horses with no or restricted access to grass or access to brown grass (e.g. during drought) are likely to benefit from vitamin E supplementation, as vitamin E levels in hay and preserved forages decrease with time.
Horses eating high fat diets should have increased amounts of vitamin E added to their diet - 100-150 mg vitamin E is suggested for every 100 ml of oil added to the diet, or supplement 2 mg/kg bodyweight.
Exercise increases production of free radicals, so levels of antioxidants including vitamin E should increase accordingly to prevent signs of oxidative damage in working horses such as sore muscles, stiffness, and slow recovery from hard work.
Natural v synthetic
In humans, according to the National Institutes of Health "One mg vitamin E (alpha-tocopherol) (natural) is equivalent to 1 mg RRR-alpha-tocopherol (natural) or 2 mg all rac-alpha-tocopherol (synthetic)."
IU and mg
From 2019 the form of vitamin E should be listed in ingredients/analyses (not just "vitamin E") as either mg or mgTE or alpha-TE (alpha-tocopherol equivalents), a term established to account for the differences in biological activity of the various forms of vitamin E.
1 mg of alpha-tocopherol = 1 TE or 1 mgTE.
1 mg of natural RRR-alpha-Tocopherol = 1 mgTE.
1 mg of synthetic All-rac-alpha-Tocopherol = 0.5 mgTE.
1 IU of natural vitamin E including ester forms (d- or RRR-alpha tocopherol, RRR-alpha-tocopheryl acetate, RRR-alpha-tocopheryl succinate) = 0.67 mg. 1 mg natural vitamin E = 1.49 IU.
1 IU of synthetic vitamin E including ester forms (dl- or all-rac-alpha tocopherol, all rac-alpha-tocopheryl acetate, all rac-alpha-tocopheryl succinate) = 0.45 mg. 1 mg synthetic vitamin E = 2.22 IU.
(Source: Nestle Quality Assurance Centre Vitamin A, E & D Unit Change)
Typical amounts of vitamin E in the diet
Fresh forage and immature forage hay/haylage can contain 30-100 IU/kg DM vitamin E. Vitamin E levels in hay and haylage can also be very low. Cereals typically contain 20-30 IU/kg DM.
Vitamin E is destroyed by oxidation, and most food processes reduce vitamin E - milling, flaking, shredding, harvesting, drying and storing hay. High temperatures and humidity exacerbate vitamin E loss.
Absorption - which form of vitamin E should be used?
Vitamin E is supplemented as alpha-tocopherol. RRR-alpha tocopheral (also called d-alpha tocopheral) is natural vitamin E. all-rec-alpha tocopheral (also called dl-alpha tocopherol) is synthetic vitamin E. Research has suggested that bioavailability (how easily the vitamin E is absorbed from the digestive tract) is lowest with synthetic forms, better with natural acetate and alcohol forms, and best with water-soluble forms like Nano-e, which has been shown to rapidly increase blood levels of vitamin E. Natural d-alpha tocopherol is unstable, and therefore esterified, i.e. joined to an acid like acetate, to protect the anti-oxidant properties of the alpha-tocopherol, and released for absorption by enzymes during digestion.
Natural vitamin E RRR-alpha tocopherol has the highest biological activity at 1.49 IU per mg, with synthetic all-rac-alpha tocopherol having 1.10 IU/mg. Supplemental vitamin E is in ester form, with RRR-alpha-tocopheral acetate having 1.36 IU per mg and all-rac-alpha tocopheral acetate 1.00 IU per mg (source: NRC Nutrient Requirements of Horses 2007 and Equine Applied and Clinical Nutrition 2013).
Vitamin E is soluble in fat and absorption may be increased by feeding vitamin E mixed with oil.
Vitamin E is influenced by selenium status, and both vitamin E and selenium should be fed in appropriate amounts.
Fagan MM, Harris P, Adams A, Pazdro R, Krotky A, Call J, Duberstein KJ.
Form of Vitamin E Supplementation Affects Oxidative and Inflammatory Response in Exercising Horses.
J Equine Vet Sci. 2020 Aug;91:103103. doi: 10.1016/j.jevs.2020.103103. Epub 2020 Apr 29. PMID: 32684249.
How are body concentrations of vitamin E tested?
As absorption and utilization of supplemental vitamin E may be individual to each horse, it is sometimes recommended to ensure that the diet provides the NRC recommended amount of vitamin E, and then test blood vitamin E concentrations to establish whether further vitamin E supplementation is required.
Vitamin E (alpha-tocopherol)status can be tested in blood serum or plasma. Normal reference range concentrations of alpha-tocopherol in horse plasma and serum are:
>2 μg/mL adequate
1.5 - 2 μg/mL marginal
<2 μg/mL deficient
However, alpha-tocopherol concentrations can fluctuate significantly, and a single serum sample may be an unsatisfactory indicator of vitamin E status.
Vitamin E in Equine Nutrition: Three Questions - Stephanie Valberg, KER September 2019
Adult horse requirement is 1-2 IU vitamin E per kg bodyweight per day. Grazing fresh grass provided 350-3000 IU of vitamin E per day to horses eating 1.7% of their bodyweight of pasture grass, depending on the season. Vitamin E degrades with feed/forage storage. Supplemented vitamin E is either natural: RRR-alpha-tocopheral or synthetic: all-rac-alpha-tocopheral or dl-alpha-tocopherol. Natural has been shown to have higher activity than synthetic.
When Do Horses Need Vitamin E? Clair Thunes, August 2019, www.thehorse.com
Vitamin E in horses - UC Davis Veterinary Medicine
Vitamin E - Oregon State University (relating to humans)
Vitamin E – An Essential Nutrient for Horses - E Kane & RL Stuart
(Author bias: note Stuart Products sell natural vitamin E)
Vitamin E and Selenium Are Powerful Protectors - Dr Eleanor Kellon July 2018
Brown JC, Valberg SJ, Hogg M, Finno CJ
Effects of feeding two RRR-α-tocopherol formulations on serum, cerebrospinal fluid and muscle α-tocopherol concentrations in horses with subclinical vitamin E deficiency
Equine Vet J. 2017;49(6):753-758. doi:10.1111/evj.12692
Duberstein KJ, Pazdro R, Lee KC, Abrams A, Kane E, Stuart RL
Effect of Supplemental Vitamin E Form on Serum α-Tocopherol Levels and Blood Oxidative Stress Parameters in Response to a Novel Exercise Challenge
JEVS October 2017 Vol 57, Pages 61–66
(Author bias: note RL Stuart of Stuart Products sells natural vitamin E).
Finno CJ, Valberg SJ.
A comparative review of vitamin E and associated equine disorders.
J Vet Intern Med. 2012 Nov-Dec;26(6):1251-66. doi: 10.1111/j.1939-1676.2012.00994.x. Epub 2012 Aug 27. PMID: 22925200.
Pusterla N, Puschner B, Steidl S, Collier J, Kane E, Stuart RL
α-Tocopherol concentrations in equine serum and cerebrospinal fluid after vitamin E supplementation
Vet Rec. 2010;166:366–368
Synthetic all-rac-α-TP acetate given at 10,000 IU/day for 14 days increased serum α-TP from deficient to normal ranges, but had no impact on cerebrospinal fluid (CSF) α-TP concentrations.
Pagan JD, Kane E, Nash D
Form and source of tocopherol affects Vitamin E status in Thoroughbred horses
Pferdeheilkunde. 2005;21:101–102
Am J Vet Res. 1992 Dec;53(12):2228-34
Variability of alpha-tocopherol values associated with procurement, storage, and freezing of equine serum and plasma samples
Craig AM, Blythe LL, Rowe KE, Lassen ED, Barrington R, Walker KC
Alpha tocopherol levels in blood samples can be reduced by haemolysis (33% decrease), leaving the blood sample in contact with the rubber stopper of the collection tube during refrigeration for 72 hours (10% decrease), and storing blood at 20-25 degrees C for 72 hours (5% decrease).
Vitamin E in horses - UC Davis Veterinary Medicine
Bampidis et al.
Assessment of a feed additive consisting of all-rac-alpha-tocopheryl acetate (vitamin E) for all animal species for the renewal of its authorisation (NHU Europe GmbH)
EFSA Journal April 2021 Vol 19, Issue 4 https://doi.org/10.2903/j.efsa.2021.6533
Vitamin K
Vitamin K acts as a cofactor for vitamin-K-dependent carboxylase, an enzyme that catalyzes the synthesis of gamma-carboxyglutamic acid (GLA) from glutamic acid. GLA is essential for blood clotting, vascular health and bone metabolism.
Vitamin K is found in green leafy plants (as phylloquinone K1), produced by bacteria in the intestinal tract, and is supplemented as synthetic K3 (which is water-soluble). All forms of vitamin K are converted to hydroquinone, the active form, in the liver.
Forages contain 3-20 mg/kg DM vitamin K, and both fresh and dried forage should meet the horse's dietary requirement for vitamin K. Cereals contain very little (0.2-0.4 mg/kg DM).
The vitamin K in forage plus the vitamin K synthesized by intestinal bacteria should meet a horse's requirements. However, only small amounts of vitamin K are stored in the body, therefore deficiency could develop, e.g. following reduced feed intake, gut disturbance (e.g. following antibiotic treatment) or impaired fat absorption.
Signs of vitamin K deficiency include decreased blood clotting ability, increased susceptibility to haemorrhage (and haematomas), and increased blood clotting time. Nosebleeds (epistaxis) may be a first sign of vitamin K deficiency, but other common reasons e.g. exercise-induced pulmonary hemorrhage, ethmoid hematoma or guttural pouch mycosis should be considered.
Vitamin K deficiency has only been documented in horses following exposure to dicoumarol (from coumarin found on mouldy clover or haylage) or warfarin (NB warfarin toxicity can be exacerbated by NSAIDs).
Supplemental vitamin K may be considered for horses with liver problems, intestinal bacterial disturbance or problems absorbing fat (give water soluble vitamin K to horses with fat malabsorption).
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
Bone Growth in Young Horses: The Importance of Vitamin K - Dr Kathleen Crandell - KER June 2015
Vitamin K is involved in blood clotting and bone metabolism.
Osteocalcin, a blood-clotting protein that is also involved in bone mineralization, may be a more sensitive marker for vitamin K status than blood coagulation factors.
"Natural sources of vitamin K in equine diets are phylloquinone (K-1), found in green leafy plants, and menaquinone (K-2), which is produced by bacteria in the digestive system. Both phylloquinone and menaquinone are converted to the active vitamin (hydroquinone) in the liver."
TLS note: When giving NSAIDs (or herbs that can increase bleeding), would it be a good idea to ensure a supplemental source of vitamin K, particularly if forage intake is reduced and/or there is a risk of gut disturbance (because the horse is taking NSAIDs or is ill)?
Vitamin K acts as a cofactor for vitamin-K-dependent carboxylase, an enzyme that catalyzes the synthesis of gamma-carboxyglutamic acid (GLA) from glutamic acid. GLA is essential for blood clotting, vascular health and bone metabolism.
Vitamin K is found in green leafy plants (as phylloquinone K1), produced by bacteria in the intestinal tract, and is supplemented as synthetic K3 (which is water-soluble). All forms of vitamin K are converted to hydroquinone, the active form, in the liver.
Forages contain 3-20 mg/kg DM vitamin K, and both fresh and dried forage should meet the horse's dietary requirement for vitamin K. Cereals contain very little (0.2-0.4 mg/kg DM).
The vitamin K in forage plus the vitamin K synthesized by intestinal bacteria should meet a horse's requirements. However, only small amounts of vitamin K are stored in the body, therefore deficiency could develop, e.g. following reduced feed intake, gut disturbance (e.g. following antibiotic treatment) or impaired fat absorption.
Signs of vitamin K deficiency include decreased blood clotting ability, increased susceptibility to haemorrhage (and haematomas), and increased blood clotting time. Nosebleeds (epistaxis) may be a first sign of vitamin K deficiency, but other common reasons e.g. exercise-induced pulmonary hemorrhage, ethmoid hematoma or guttural pouch mycosis should be considered.
Vitamin K deficiency has only been documented in horses following exposure to dicoumarol (from coumarin found on mouldy clover or haylage) or warfarin (NB warfarin toxicity can be exacerbated by NSAIDs).
Supplemental vitamin K may be considered for horses with liver problems, intestinal bacterial disturbance or problems absorbing fat (give water soluble vitamin K to horses with fat malabsorption).
Feeding Your Horse Properly, Part 4 The Fat Soluble Vitamins (A, D, E, K) - Dr. Eleanor Kellon, VMD
Bone Growth in Young Horses: The Importance of Vitamin K - Dr Kathleen Crandell - KER June 2015
Vitamin K is involved in blood clotting and bone metabolism.
Osteocalcin, a blood-clotting protein that is also involved in bone mineralization, may be a more sensitive marker for vitamin K status than blood coagulation factors.
"Natural sources of vitamin K in equine diets are phylloquinone (K-1), found in green leafy plants, and menaquinone (K-2), which is produced by bacteria in the digestive system. Both phylloquinone and menaquinone are converted to the active vitamin (hydroquinone) in the liver."
TLS note: When giving NSAIDs (or herbs that can increase bleeding), would it be a good idea to ensure a supplemental source of vitamin K, particularly if forage intake is reduced and/or there is a risk of gut disturbance (because the horse is taking NSAIDs or is ill)?
Volatile Fatty Acids (VFAs - also SCFAs)
Research measuring VFAs:
Animal. 2017 Jul;11(7):1146-1152. doi: 10.1017/S1751731116002603. Epub 2016 Dec 8.
Effect of concentrate feeding sequence on equine hindgut fermentation parameters.
Sadet-Bourgeteau S, Philippeau C, Julliand V
Research measuring VFAs:
Animal. 2017 Jul;11(7):1146-1152. doi: 10.1017/S1751731116002603. Epub 2016 Dec 8.
Effect of concentrate feeding sequence on equine hindgut fermentation parameters.
Sadet-Bourgeteau S, Philippeau C, Julliand V
Water
Horses take in water by drinking, and also in their feed - grass is often around 80% water. Horses lose water from their body in urine, dung, sweat and a small amount in their breath.
Soaking and steaming hay are good ways to increase water intake, as is feeding a wet warm mash based on feeds like sugar beet/beet pulp or soya hulls.
For correct hydration, horses need electrolytes as well as water. Most electrolytes are provided by a normal forage diet, but sodium is low in forage and should be provided to the horse, in bucket feed and/or as a plain salt lick.
Horses take in water by drinking, and also in their feed - grass is often around 80% water. Horses lose water from their body in urine, dung, sweat and a small amount in their breath.
Soaking and steaming hay are good ways to increase water intake, as is feeding a wet warm mash based on feeds like sugar beet/beet pulp or soya hulls.
For correct hydration, horses need electrolytes as well as water. Most electrolytes are provided by a normal forage diet, but sodium is low in forage and should be provided to the horse, in bucket feed and/or as a plain salt lick.
Whey protein
Feeding horses whey protein - KER April 2016
Whey protein is considered a high quality protein source and is suitable for consumption by horses. Over 60% of the proteins in whey are essential amino acids, and whey protein is particularly high in branch chain amino acids (BCAA), which are particularly important for muscle repair following exercise. Whey protein isolate is the best form for horses due to its low lactose content. Whey is a source of the protein lactoferrin, which has been found to have a growth simulating effect on bone, and may help reduce the mineral loss in bones that can be caused by long-term confinement.
Milk Protein - www.milkfacts.info
Milk contains all the essential amino acids required by mammals. 3.3% of cow's milk is protein, 18% of which is whey protein, and the other 82% casein. Whey protein contains a large amount of sulphur-containing amino acids, and doesn't contain phosphorus. Whey protein consists of ß-lactoglobulin, α-lactalbumin, blood serum albumin, immunoglobulins, lactoferrin, transferrin, and minor proteins and enzymes. "The function of ß-lactoglobulin is thought to be a carrier of vitamin A." "α-Lactalbumin plays a critical role in the synthesis of lactose in the mammary gland. Immunoglobulins play a role in the animal's immune system.... Lactoferrin and transferrin play an important role in iron absorption".
Feeding horses whey protein - KER April 2016
Whey protein is considered a high quality protein source and is suitable for consumption by horses. Over 60% of the proteins in whey are essential amino acids, and whey protein is particularly high in branch chain amino acids (BCAA), which are particularly important for muscle repair following exercise. Whey protein isolate is the best form for horses due to its low lactose content. Whey is a source of the protein lactoferrin, which has been found to have a growth simulating effect on bone, and may help reduce the mineral loss in bones that can be caused by long-term confinement.
Milk Protein - www.milkfacts.info
Milk contains all the essential amino acids required by mammals. 3.3% of cow's milk is protein, 18% of which is whey protein, and the other 82% casein. Whey protein contains a large amount of sulphur-containing amino acids, and doesn't contain phosphorus. Whey protein consists of ß-lactoglobulin, α-lactalbumin, blood serum albumin, immunoglobulins, lactoferrin, transferrin, and minor proteins and enzymes. "The function of ß-lactoglobulin is thought to be a carrier of vitamin A." "α-Lactalbumin plays a critical role in the synthesis of lactose in the mammary gland. Immunoglobulins play a role in the animal's immune system.... Lactoferrin and transferrin play an important role in iron absorption".
Willow / white willow (Salix alba) / salicin/salicylic acid
Can horses eat willow trees? Are there any health benefits?
Willow (Salix species) does not appear to be poisonous for horses, and research has suggested large amounts of willow are safe for rabbits to eat.
Fresh leaves may contain varying levels of protein and carbohydrates. Tannins and other metabolites may reduce the amount of protein available to an animal ingesting willow.
Willow bark contains salicin, which is converted into salicylic acid (similar to aspirin) in the body, and may have analgesic, anti-inflammatory and antipyretic effects, although the amount of salicylic acid produced from salicin is small. Side effects associated with ingestion of willow bark in humans include allergic reactions and gastro-intestinal symptoms.
Nutritional content
Thin willow stems with leaves were found to contain an average of 3.5% WSC (on a dry matter basis) in the UK at the end of June by Smith et al. 2014; Kemp et al. 2003 suggest WSC of 12.3% DM for willow.
Martin, Benjamin
The Suitability of Willow Trees as Animal Forage and their Application in Zoological Institutions
2019 MSc thesis University of Guelph
Willow has been used as a drought feed supplement and protein source.
5 willow cultivars were sampled once a month in June, July, August and September, around 13:00 hours, in Toronto - branches were cut 15 cm from the basal attachment point of the tree trunk. The samples were separated into leaf, bark and twig material.
The author warns that when using crude protein values from tree material, tannins and other protein-binding metabolites can significantly reduce the quantities of proteins and amino acids available to the animal.
In June and July (Canada) leaf dry matter content averaged 39.5%, crude protein 14.5%, ADF 25.1%, NDF 35.4% (all figures DM).
In August and September leaf dry matter content averaged 43.7%, CP 14.5%, ADF 25%, NDF 35.3%.
Willow trees are known for their analgesic, anti-inflammatory and antipyretic effects, thought to be due to the salicylic glycoside salicin and its derivatives, but a variety of compounds may work synergistically to produce anti-inflammatory effects.
36 young rabbits were divided into 4 groups:
Control C100 - ad lib rabbit feed pellets only
Control C50 - restricted rabbit feed pellets only
TL - restricted rabbit feed pellets plus ad lib fresh willow leaf material (from Salix miyabeana)
TB - restricted rabbit feed pellets plus ad lib willow bark material (from Salix miyabeana).
All groups grew well, maintained blood chemistry values within reference ranges and did not show significant changes in markers of systemic inflammation. There was no evidence that eating fresh willow changed blood clotting, and there were no indications of toxicity or cellular damage, suggesting that high levels of willow ingestion appers to be a safe and effective forage material for rabbits.
Smith J, Juoppala K, Yanez-Ruiz D, Leach K, Rinne M
Nutritional and fermentation quality of ensiled willow from an integrated feed and bioenergy agroforestry system in UK
No. 30 (2014): Days of Agricultural Science 2014
Samples of first year regrowth Salix viminalis willow branches with a stem diameter < 8 mm, and leaves only, were picked at the end of June and analysed in Finland.
On a dry matter basis, the leaf on stem samples contained:
26.5% dry matter
7.1% ash
16.7% crude protein
3.53% WSC
57.3% NDF
41.0% ADF
18.4% lignin
Unfortunately the leaves only were not analysed as raw material, only after being ensiled (fermented to silage). After 6 months ensiling, on a dry matter basis the leaf only ensiled samples contained:
28.2% dry matter
9.4% ash
21.9% crude protein
1.54% WSC
28.7% NDF
19.9% ADF
5.2% lignin
WSC would be expected to decrease with ensiling (WSC is converted to fermentation acids (VFAs)).
Tree fodder nutritive value and digestibility peaks in spring and decreases through to autumn.
Tannins were almost twice as high in the leaf only silage compared to the leaf and stem silage.
Referencing Tolera et al. 1997, the authors suggest that tree fodder is generally higher in protein and minerals than dry season pasture, but the prescence of tannins and other compounds may reduce digestibility, availability of protein, palatability and intake.
Kemp PD, Barry TN, Douglas GB
Edible forage yield and nutritive value of poplar and willow
New Zealand Grassland Association Workshop Proceedings 2003
Edible forage (leaves plus stems of <5 mm) from Tangoio willow in New Zealand was picked in January and February for 3 years and analysed using "standard laboratory methods" at Massey University and AgReseaarch Grasslands laboratory.
Measured in December and March/spring and summer, willow mean crude protein ranged from 7.1 to 17.1% DM and ME from 9.6 to 11.6 MJ/kg DM.
WSC for willow is given as 12.3% DM, NDF 38.1%, ADF 26.4%, lignin 13.4%, salicin 0.5%, condensed tannins 5.2%.
According to the authors, senesced or dead leaves will have a low nutritive value with an ME of 7-8 MJ/kg DM.
Anti-inflammatory and pain relieving properties
Willow bark contains salicin, which is converted into salicylic acid in the body. Salicylic acid may work similarly to aspirin by blocking cyclo-oxygenase enzymes that are involved in inflammation and pain.
The amount of salicylic acid produced from salicin is small.
Side effects associated with willow bark medicines (in humans) include allergic reactions such as rash, itching, asthma, and stomach/intestine symptoms such as nausea, vomiting, abdominal pain, diarrhoea. Salicin should not be taken by people with asthma, stomach ulcers, liver or kidney dysfunction, clotting disorders, or by pregnant women or children.
Source:
Salicis cortex - Committee on Herbal Medicinal Products, European Medicines Agency, 2017
Vlachojannis J, Magora F, Chrubasik S.
Willow species and aspirin: different mechanism of actions.
Phytother Res. 2011 Jul;25(7):1102-4. doi: 10.1002/ptr.3386. PMID: 21226125.
"Willow species contain only a low quantity of the prodrug salicin which is metabolized during absorption into various salicylate derivatives. If calculated as salicylic acid, the daily salicin dose is insufficient to produce analgesia. Salicylic acid concentrations following an analgesic dose of aspirin are an order of magnitude higher. Flavonoids and polyphenols contribute to the potent willow bark analgesic and anti-inflammatory effect. The multi-component active principle of willow bark provides a broader mechanism of action than aspirin and is devoid of serious adverse events. In contrast to synthetic aspirin, willow bark does not damage the gastrointestinal mucosa. An extract dose with 240 mg salicin had no major impact on blood clotting. In patients with known aspirin allergy willow bark products are contraindicated."
The Curious History of Herbs and Plants as Medicines - Dr David Ramey, February 2018
Dr Ramey writes that aspirin (acetylsalicylic acid) is a derivative of salicylic acid, which is found in the bark and leaves of white willow (Salix alba) as salicin. 14 grams of willow bark contains around 1 gram of salicin (which is about half as potent as aspirin), but willow bark contains tannins, and both the tannins and salicin are irritating to the stomach and likely to cause digestive problems.
Aspirin (acetylsalicylic acid) is not recommended for horses with laminitis because of the increased risk of bleeding and other potential side effects (including gastro-intestinal irritation, liver and kidney damage and protein loss), and its weak anti-inflammatory and pain relieving properties in horses. See www.thelaminitissite - Aspirin.
Can horses eat willow trees? Are there any health benefits?
Willow (Salix species) does not appear to be poisonous for horses, and research has suggested large amounts of willow are safe for rabbits to eat.
Fresh leaves may contain varying levels of protein and carbohydrates. Tannins and other metabolites may reduce the amount of protein available to an animal ingesting willow.
Willow bark contains salicin, which is converted into salicylic acid (similar to aspirin) in the body, and may have analgesic, anti-inflammatory and antipyretic effects, although the amount of salicylic acid produced from salicin is small. Side effects associated with ingestion of willow bark in humans include allergic reactions and gastro-intestinal symptoms.
Nutritional content
Thin willow stems with leaves were found to contain an average of 3.5% WSC (on a dry matter basis) in the UK at the end of June by Smith et al. 2014; Kemp et al. 2003 suggest WSC of 12.3% DM for willow.
Martin, Benjamin
The Suitability of Willow Trees as Animal Forage and their Application in Zoological Institutions
2019 MSc thesis University of Guelph
Willow has been used as a drought feed supplement and protein source.
5 willow cultivars were sampled once a month in June, July, August and September, around 13:00 hours, in Toronto - branches were cut 15 cm from the basal attachment point of the tree trunk. The samples were separated into leaf, bark and twig material.
The author warns that when using crude protein values from tree material, tannins and other protein-binding metabolites can significantly reduce the quantities of proteins and amino acids available to the animal.
In June and July (Canada) leaf dry matter content averaged 39.5%, crude protein 14.5%, ADF 25.1%, NDF 35.4% (all figures DM).
In August and September leaf dry matter content averaged 43.7%, CP 14.5%, ADF 25%, NDF 35.3%.
Willow trees are known for their analgesic, anti-inflammatory and antipyretic effects, thought to be due to the salicylic glycoside salicin and its derivatives, but a variety of compounds may work synergistically to produce anti-inflammatory effects.
36 young rabbits were divided into 4 groups:
Control C100 - ad lib rabbit feed pellets only
Control C50 - restricted rabbit feed pellets only
TL - restricted rabbit feed pellets plus ad lib fresh willow leaf material (from Salix miyabeana)
TB - restricted rabbit feed pellets plus ad lib willow bark material (from Salix miyabeana).
All groups grew well, maintained blood chemistry values within reference ranges and did not show significant changes in markers of systemic inflammation. There was no evidence that eating fresh willow changed blood clotting, and there were no indications of toxicity or cellular damage, suggesting that high levels of willow ingestion appers to be a safe and effective forage material for rabbits.
Smith J, Juoppala K, Yanez-Ruiz D, Leach K, Rinne M
Nutritional and fermentation quality of ensiled willow from an integrated feed and bioenergy agroforestry system in UK
No. 30 (2014): Days of Agricultural Science 2014
Samples of first year regrowth Salix viminalis willow branches with a stem diameter < 8 mm, and leaves only, were picked at the end of June and analysed in Finland.
On a dry matter basis, the leaf on stem samples contained:
26.5% dry matter
7.1% ash
16.7% crude protein
3.53% WSC
57.3% NDF
41.0% ADF
18.4% lignin
Unfortunately the leaves only were not analysed as raw material, only after being ensiled (fermented to silage). After 6 months ensiling, on a dry matter basis the leaf only ensiled samples contained:
28.2% dry matter
9.4% ash
21.9% crude protein
1.54% WSC
28.7% NDF
19.9% ADF
5.2% lignin
WSC would be expected to decrease with ensiling (WSC is converted to fermentation acids (VFAs)).
Tree fodder nutritive value and digestibility peaks in spring and decreases through to autumn.
Tannins were almost twice as high in the leaf only silage compared to the leaf and stem silage.
Referencing Tolera et al. 1997, the authors suggest that tree fodder is generally higher in protein and minerals than dry season pasture, but the prescence of tannins and other compounds may reduce digestibility, availability of protein, palatability and intake.
Kemp PD, Barry TN, Douglas GB
Edible forage yield and nutritive value of poplar and willow
New Zealand Grassland Association Workshop Proceedings 2003
Edible forage (leaves plus stems of <5 mm) from Tangoio willow in New Zealand was picked in January and February for 3 years and analysed using "standard laboratory methods" at Massey University and AgReseaarch Grasslands laboratory.
Measured in December and March/spring and summer, willow mean crude protein ranged from 7.1 to 17.1% DM and ME from 9.6 to 11.6 MJ/kg DM.
WSC for willow is given as 12.3% DM, NDF 38.1%, ADF 26.4%, lignin 13.4%, salicin 0.5%, condensed tannins 5.2%.
According to the authors, senesced or dead leaves will have a low nutritive value with an ME of 7-8 MJ/kg DM.
Anti-inflammatory and pain relieving properties
Willow bark contains salicin, which is converted into salicylic acid in the body. Salicylic acid may work similarly to aspirin by blocking cyclo-oxygenase enzymes that are involved in inflammation and pain.
The amount of salicylic acid produced from salicin is small.
Side effects associated with willow bark medicines (in humans) include allergic reactions such as rash, itching, asthma, and stomach/intestine symptoms such as nausea, vomiting, abdominal pain, diarrhoea. Salicin should not be taken by people with asthma, stomach ulcers, liver or kidney dysfunction, clotting disorders, or by pregnant women or children.
Source:
Salicis cortex - Committee on Herbal Medicinal Products, European Medicines Agency, 2017
Vlachojannis J, Magora F, Chrubasik S.
Willow species and aspirin: different mechanism of actions.
Phytother Res. 2011 Jul;25(7):1102-4. doi: 10.1002/ptr.3386. PMID: 21226125.
"Willow species contain only a low quantity of the prodrug salicin which is metabolized during absorption into various salicylate derivatives. If calculated as salicylic acid, the daily salicin dose is insufficient to produce analgesia. Salicylic acid concentrations following an analgesic dose of aspirin are an order of magnitude higher. Flavonoids and polyphenols contribute to the potent willow bark analgesic and anti-inflammatory effect. The multi-component active principle of willow bark provides a broader mechanism of action than aspirin and is devoid of serious adverse events. In contrast to synthetic aspirin, willow bark does not damage the gastrointestinal mucosa. An extract dose with 240 mg salicin had no major impact on blood clotting. In patients with known aspirin allergy willow bark products are contraindicated."
The Curious History of Herbs and Plants as Medicines - Dr David Ramey, February 2018
Dr Ramey writes that aspirin (acetylsalicylic acid) is a derivative of salicylic acid, which is found in the bark and leaves of white willow (Salix alba) as salicin. 14 grams of willow bark contains around 1 gram of salicin (which is about half as potent as aspirin), but willow bark contains tannins, and both the tannins and salicin are irritating to the stomach and likely to cause digestive problems.
Aspirin (acetylsalicylic acid) is not recommended for horses with laminitis because of the increased risk of bleeding and other potential side effects (including gastro-intestinal irritation, liver and kidney damage and protein loss), and its weak anti-inflammatory and pain relieving properties in horses. See www.thelaminitissite - Aspirin.
Wood chewing
Horses evolved to spend the majority of their day grazing and browsing on high fibre, low energy plants. Wood chewing may be associated with a need to graze/feed, and/or with a need for more fibre in the diet, as increasing the fibre content of the diet will often reduce wood chewing. Willard et al. 1997 found that horses eating a concentrate only diet chewed wood significantly more than horses eating a hay diet.
Wood chewing can be considered an abnormal behaviour that may be associated with diet and feed management. Waters et al. 2002 found that wood chewing often preceded crib-biting in young horses.
Waters AJ, Nicol CJ, French NP
Factors influencing the development of stereotypic and redirected behaviours in young horses: findings of a four year prospective epidemiological study
Equine Vet J. 2002 Sep;34(6):572-9
Redbo I, Redbo-Torstensson P, Ödberg FO, Hedendahl A
Factors affecting behavioural disturbances in race-horses
Animal Science Volume 66, Issue 2 April 1998, pp. 475-481
Willard JG, Willard JC, Wolfram SA, Baker JP.
Effect of diet on cecal pH and feeding behavior of horses.
J Anim Sci. 1977 Jul;45(1):87-93.
Horses evolved to spend the majority of their day grazing and browsing on high fibre, low energy plants. Wood chewing may be associated with a need to graze/feed, and/or with a need for more fibre in the diet, as increasing the fibre content of the diet will often reduce wood chewing. Willard et al. 1997 found that horses eating a concentrate only diet chewed wood significantly more than horses eating a hay diet.
Wood chewing can be considered an abnormal behaviour that may be associated with diet and feed management. Waters et al. 2002 found that wood chewing often preceded crib-biting in young horses.
Waters AJ, Nicol CJ, French NP
Factors influencing the development of stereotypic and redirected behaviours in young horses: findings of a four year prospective epidemiological study
Equine Vet J. 2002 Sep;34(6):572-9
Redbo I, Redbo-Torstensson P, Ödberg FO, Hedendahl A
Factors affecting behavioural disturbances in race-horses
Animal Science Volume 66, Issue 2 April 1998, pp. 475-481
Willard JG, Willard JC, Wolfram SA, Baker JP.
Effect of diet on cecal pH and feeding behavior of horses.
J Anim Sci. 1977 Jul;45(1):87-93.
Yea-sacc
See also Probiotics and Postbiotics (SCFP).
Saccharomyces cerevisiae (S. cerevisiae) fermentation products (SCFP), commonly called yea-sacc, are used pre- and postbiotic feed supplements for horses and other animals.
Probiotics have been defined as "a live microbial feed supplement which beneficially affects the host animal by improving its intestinal microbial balance”, and
postbiotics as a “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host."
Perricone V, Sandrini S, Irshad N, Comi M, Lecchi C, Savoini G, Agazzi A.
The Role of Yeast Saccharomyces cerevisiae in Supporting Gut Health in Horses: An Updated Review on Its Effects on Digestibility and Intestinal and Fecal Microbiota.
Animals (Basel). 2022 Dec 9;12(24):3475. doi: 10.3390/ani12243475. PMID: 36552396; PMCID: PMC9774806.
This review concludes that "while the addition of S. cerevisiae to the diet of horses could somehow benefit gut health, e.g., improving, in some cases, the nutrient digestibility or modulating fibrolytic and amylolytic bacteria, its efficacy and results are quite contradictory and highly variable."
How diet can affect the equine microbiome - J Murray, BEVA September 2022
At BEVA 2022, J Murray reported:
Probiotics are "live micro-organisms which, when administered in adequate amounts, confer a health benefit on the host". No probiotics are registered for use in horses in the EU. 3 strains of yea-sacc (Saccharomyces cerevisiae) are registered for use in horses in the EU.
Research has shown that feeding yeast (yea-sacc) may increase fibre digestibility, encourage fibre-degrading microbes and reduce lactic-acid producing bacteria, all of which are good for gut health. Yeast may also help keep the gut healthy if starch is fed or horses are stressed e.g. from travelling. Yeast needs to be fed daily.
To maintain a stable microbial population:
- feed a high fibre diet
- keep starch to a minimum, if starch must be fed avoid large meals - feed little amounts more frequently
- make all changes to the diet gradually, over at least 2 weeks, including changing between pasture/fresh grass and preserved forage
- consider using a yeast supplement
[TLS Note: Jo-Anne Murray is a consultant for supplement company Premier Performance].
Garber A, Hastie PM, Farci V, Bulmer L, Alzahal O, Murray JMD.
The effect of supplementing pony diets with yeast on 1. In vivo and in vitro digestibility, faecal pH and particle size.
Animal. 2020 Dec;14(12):2481-2492. doi: 10.1017/S1751731120001524. Epub 2020 Jun 25. PMID: 32580794.
6 ponies were fed each of a high fibre and high starch diet with and without a supplement containing 2 g yea-sacc per day, for 19 days.
Faecal pH was the same whether yeast was added or not for the high fibre diet. Faecal pH was lower for high starch than high fibre diets, but pH was lower for high starch no yeast than high starch plus yeast. Yeast supplementation slightly increased apparent digestion of fibre on the high fibre diet, and more significantly increased apparent digestion of fibre on the high starch diet.
Garber A, Hastie PM, Farci V, McGuinness D, Bulmer L, Alzahal O, Murray JMD.
The effect of supplementing pony diets with yeast on 2. The faecal microbiome.
Animal. 2020 Dec;14(12):2493-2502. doi: 10.1017/S1751731120001512. Epub 2020 Jun 25. PMID: 32580805.
6 ponies were fed each of a high fibre and high starch diet with and without a supplement containing 2 g yea-sacc per day, for 19 days. Feeding yea-sacc altered the faecal microbial population of the ponies. Bacteria that produce VFAs were increased, therefore "it may be suggested that S. cerevisiae may have a positive effect on the utilisation of fibre in ponies by increasing the number of fibrolytic bacteria and thus increasing the nutritive value of the diet for the horses" [which is something that may not be advantageous in horses that need to lose weight!]. Supplementing yea-sacc to the high starch diet decreased lactate producing bacteria and increased gut pH, in comparison to the high starch diet without yeast, suggesting that yeast supplementation may help to maintain less acidic GI environments and possibly reduce the negative effects of high starch diets.
Conclusions:
Yeast supplementation altered the faecal microbial population of ponies.
Previous research showed that yeast supplementation increased total tract apparent nutrient digestibility of high fibre and high starch diets.
Further research is needed e.g. to quantify VFA production with microbiota changes.
Equine Applied and Clinical Nutrition (2013) says "there has been no evidence to prove that providing probiotics to animals with healthy thriving GI flora will have any beneficial effect". Basically, there are some studies that suggest that feeding yea-sacc (or other Saccharomyces species) to horses that are on high cereal/concentrate diets may increase the pH of the hind gut and improve fibre digestion, but "it is hard to make a recommendation for its use in aiding fiber digestion in horses provided typical forage based rations".
The NRC Nutrient Requirements of Horses 2007 goes into a bit more detail, but still suggests that minimal effects have been seen when adding live yeast to the diet except when high starch/low forage diets are fed. Some research has suggested increased protein digestibility with live yeast - but this appears to be the difference between protein ingested and excreted, it would presumably be protein "digestion" in the hind gut (rather than the small intestine) that increased, and it is currently not known whether horses can benefit from protein once it reaches the hind gut - it may feed the microorganisms, but have no benefit to the horse - so any increase in protein digestibility recorded by this intake minus excretion method may have no benefit for the horse - see
Morgan LM, Coverdale JA, Froetschel, Yoon I
Effect of Yeast Culture Supplementation on Digestibility of Varying Forage Quality in Mature Horses
Journal of Equine Veterinary Science - Volume 27, Issue 6, June 2007, Pages 260–265
(note that the horses were fed less than 2% of their bodyweight, with 25% of their diet as concentrate, and that they went ~19 hours a day without food - so hopefully this research bears little relation to how most owners keep their horses. Note also that one of the authors works for a company producing the yeast used! - this may not be unbiased research!).
Similar research has suggested a slight increase in magnesium and phosphorus uptake, but again, looking at Pagan's paper that showed an increase in phosphorus digestibility from 22% to 27% and 19% to 24%, the (very few) horses were fed 1/3 of their diet as concentrate, fed less than 2% bodyweight in total, and probably did not have food in front of them for the majority of the day (potentially leading to increasingly acidic conditions in their GI tract), so this research may not be particularly relevant to trickle-fed horses. And again, the research is carried out by a company selling yeast.
Pagan JD
Effect of Yeast Culture Supplementation on Nutrient Digestibility in Mature Horses
Abstract 340, Journal of Animal Science, Vol. 68, Supp. 1, 1990
Murray JAMD, Brown S, O'Shaughnessy PJ, Monteiro A, Warren H, Hastie P
Effect of live yeast culture supplementation on fibrolytic and saccharolytic bacterial populations in the faeces of horses fed a high-fibre or high-starch diet
Journal of Equine Veterinary Science published online Dec 2016
Live yeast (Saccharomyces cerevisiae) was given to 4 horses fed high starch and high fibre (low starch) diets in a 4 x 4 Latin square design study (high fibre no yeast HF, high fibre + yeast HFY, high starch no yeast HS, high starch + yeast HSY). Each feeding regime lasted 31 days, with fresh faecal samples being collected on the last 3 days of each regime and analysed for bacterial content, particularly cellulolytic (Fibrobacter succinogenes and Ruminococcus flavefaciens) and saccharolytic (Streptococcus equinus and Streptococcus bovis) bacteria.
F. succinogenes was the most abundent bacteria. The high starch + yeast diet resulted in a significant reduction in relative levels of this bacterium.
R. flavefaciens was not significantly affected by diet.
S. equinus was not significantly affected by diet, although increased levels were seen in horses eating the high starch diets.
S. bovis was increased in horses eating the high starch diets.
Significant variation in levels of S. equinus and S. bovis was seen between individual horses.
Summary: the effect of live yeast on large intestinal bacterial populations was examined in horses fed a low or high starch diet. Bacterial populations were affected by diet, but not by yeast supplementation.
Yeast Cultures - Dr Bob Coleman - Alberta Agriculture Food and Rural Development
Mackenthun E, Coenen M, Vervuert I.
Effects of Saccharomyces cerevisiae supplementation on apparent total tract digestibility of nutrients and fermentation profile in healthy horses.
J Anim Physiol Anim Nutr (Berl). 2013 May;97 Suppl 1:115-20. doi: 10.1111/jpn.12043. PMID: 23639024.
Supplementing healthy horses eating cracked corn and hay with Saccharomyces cerevisiae made no significant difference to total tract digestibility of crude fibre, short-chain fatty acids, lactic acids and pH in collected dung.
See also Probiotics and Postbiotics (SCFP).
Saccharomyces cerevisiae (S. cerevisiae) fermentation products (SCFP), commonly called yea-sacc, are used pre- and postbiotic feed supplements for horses and other animals.
Probiotics have been defined as "a live microbial feed supplement which beneficially affects the host animal by improving its intestinal microbial balance”, and
postbiotics as a “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host."
Perricone V, Sandrini S, Irshad N, Comi M, Lecchi C, Savoini G, Agazzi A.
The Role of Yeast Saccharomyces cerevisiae in Supporting Gut Health in Horses: An Updated Review on Its Effects on Digestibility and Intestinal and Fecal Microbiota.
Animals (Basel). 2022 Dec 9;12(24):3475. doi: 10.3390/ani12243475. PMID: 36552396; PMCID: PMC9774806.
This review concludes that "while the addition of S. cerevisiae to the diet of horses could somehow benefit gut health, e.g., improving, in some cases, the nutrient digestibility or modulating fibrolytic and amylolytic bacteria, its efficacy and results are quite contradictory and highly variable."
How diet can affect the equine microbiome - J Murray, BEVA September 2022
At BEVA 2022, J Murray reported:
Probiotics are "live micro-organisms which, when administered in adequate amounts, confer a health benefit on the host". No probiotics are registered for use in horses in the EU. 3 strains of yea-sacc (Saccharomyces cerevisiae) are registered for use in horses in the EU.
Research has shown that feeding yeast (yea-sacc) may increase fibre digestibility, encourage fibre-degrading microbes and reduce lactic-acid producing bacteria, all of which are good for gut health. Yeast may also help keep the gut healthy if starch is fed or horses are stressed e.g. from travelling. Yeast needs to be fed daily.
To maintain a stable microbial population:
- feed a high fibre diet
- keep starch to a minimum, if starch must be fed avoid large meals - feed little amounts more frequently
- make all changes to the diet gradually, over at least 2 weeks, including changing between pasture/fresh grass and preserved forage
- consider using a yeast supplement
[TLS Note: Jo-Anne Murray is a consultant for supplement company Premier Performance].
Garber A, Hastie PM, Farci V, Bulmer L, Alzahal O, Murray JMD.
The effect of supplementing pony diets with yeast on 1. In vivo and in vitro digestibility, faecal pH and particle size.
Animal. 2020 Dec;14(12):2481-2492. doi: 10.1017/S1751731120001524. Epub 2020 Jun 25. PMID: 32580794.
6 ponies were fed each of a high fibre and high starch diet with and without a supplement containing 2 g yea-sacc per day, for 19 days.
Faecal pH was the same whether yeast was added or not for the high fibre diet. Faecal pH was lower for high starch than high fibre diets, but pH was lower for high starch no yeast than high starch plus yeast. Yeast supplementation slightly increased apparent digestion of fibre on the high fibre diet, and more significantly increased apparent digestion of fibre on the high starch diet.
Garber A, Hastie PM, Farci V, McGuinness D, Bulmer L, Alzahal O, Murray JMD.
The effect of supplementing pony diets with yeast on 2. The faecal microbiome.
Animal. 2020 Dec;14(12):2493-2502. doi: 10.1017/S1751731120001512. Epub 2020 Jun 25. PMID: 32580805.
6 ponies were fed each of a high fibre and high starch diet with and without a supplement containing 2 g yea-sacc per day, for 19 days. Feeding yea-sacc altered the faecal microbial population of the ponies. Bacteria that produce VFAs were increased, therefore "it may be suggested that S. cerevisiae may have a positive effect on the utilisation of fibre in ponies by increasing the number of fibrolytic bacteria and thus increasing the nutritive value of the diet for the horses" [which is something that may not be advantageous in horses that need to lose weight!]. Supplementing yea-sacc to the high starch diet decreased lactate producing bacteria and increased gut pH, in comparison to the high starch diet without yeast, suggesting that yeast supplementation may help to maintain less acidic GI environments and possibly reduce the negative effects of high starch diets.
Conclusions:
Yeast supplementation altered the faecal microbial population of ponies.
Previous research showed that yeast supplementation increased total tract apparent nutrient digestibility of high fibre and high starch diets.
Further research is needed e.g. to quantify VFA production with microbiota changes.
Equine Applied and Clinical Nutrition (2013) says "there has been no evidence to prove that providing probiotics to animals with healthy thriving GI flora will have any beneficial effect". Basically, there are some studies that suggest that feeding yea-sacc (or other Saccharomyces species) to horses that are on high cereal/concentrate diets may increase the pH of the hind gut and improve fibre digestion, but "it is hard to make a recommendation for its use in aiding fiber digestion in horses provided typical forage based rations".
The NRC Nutrient Requirements of Horses 2007 goes into a bit more detail, but still suggests that minimal effects have been seen when adding live yeast to the diet except when high starch/low forage diets are fed. Some research has suggested increased protein digestibility with live yeast - but this appears to be the difference between protein ingested and excreted, it would presumably be protein "digestion" in the hind gut (rather than the small intestine) that increased, and it is currently not known whether horses can benefit from protein once it reaches the hind gut - it may feed the microorganisms, but have no benefit to the horse - so any increase in protein digestibility recorded by this intake minus excretion method may have no benefit for the horse - see
Morgan LM, Coverdale JA, Froetschel, Yoon I
Effect of Yeast Culture Supplementation on Digestibility of Varying Forage Quality in Mature Horses
Journal of Equine Veterinary Science - Volume 27, Issue 6, June 2007, Pages 260–265
(note that the horses were fed less than 2% of their bodyweight, with 25% of their diet as concentrate, and that they went ~19 hours a day without food - so hopefully this research bears little relation to how most owners keep their horses. Note also that one of the authors works for a company producing the yeast used! - this may not be unbiased research!).
Similar research has suggested a slight increase in magnesium and phosphorus uptake, but again, looking at Pagan's paper that showed an increase in phosphorus digestibility from 22% to 27% and 19% to 24%, the (very few) horses were fed 1/3 of their diet as concentrate, fed less than 2% bodyweight in total, and probably did not have food in front of them for the majority of the day (potentially leading to increasingly acidic conditions in their GI tract), so this research may not be particularly relevant to trickle-fed horses. And again, the research is carried out by a company selling yeast.
Pagan JD
Effect of Yeast Culture Supplementation on Nutrient Digestibility in Mature Horses
Abstract 340, Journal of Animal Science, Vol. 68, Supp. 1, 1990
Murray JAMD, Brown S, O'Shaughnessy PJ, Monteiro A, Warren H, Hastie P
Effect of live yeast culture supplementation on fibrolytic and saccharolytic bacterial populations in the faeces of horses fed a high-fibre or high-starch diet
Journal of Equine Veterinary Science published online Dec 2016
Live yeast (Saccharomyces cerevisiae) was given to 4 horses fed high starch and high fibre (low starch) diets in a 4 x 4 Latin square design study (high fibre no yeast HF, high fibre + yeast HFY, high starch no yeast HS, high starch + yeast HSY). Each feeding regime lasted 31 days, with fresh faecal samples being collected on the last 3 days of each regime and analysed for bacterial content, particularly cellulolytic (Fibrobacter succinogenes and Ruminococcus flavefaciens) and saccharolytic (Streptococcus equinus and Streptococcus bovis) bacteria.
F. succinogenes was the most abundent bacteria. The high starch + yeast diet resulted in a significant reduction in relative levels of this bacterium.
R. flavefaciens was not significantly affected by diet.
S. equinus was not significantly affected by diet, although increased levels were seen in horses eating the high starch diets.
S. bovis was increased in horses eating the high starch diets.
Significant variation in levels of S. equinus and S. bovis was seen between individual horses.
Summary: the effect of live yeast on large intestinal bacterial populations was examined in horses fed a low or high starch diet. Bacterial populations were affected by diet, but not by yeast supplementation.
Yeast Cultures - Dr Bob Coleman - Alberta Agriculture Food and Rural Development
Mackenthun E, Coenen M, Vervuert I.
Effects of Saccharomyces cerevisiae supplementation on apparent total tract digestibility of nutrients and fermentation profile in healthy horses.
J Anim Physiol Anim Nutr (Berl). 2013 May;97 Suppl 1:115-20. doi: 10.1111/jpn.12043. PMID: 23639024.
Supplementing healthy horses eating cracked corn and hay with Saccharomyces cerevisiae made no significant difference to total tract digestibility of crude fibre, short-chain fatty acids, lactic acids and pH in collected dung.
Zeolite
Zeolite is hydrated alkali aluminum silicate.
Zeolite is toxic when inhaled.
Zeolite - www.mskcc.org
NRC Nutrient Requirements of Horses 2007 p 186:
Zeolites are potential binders of cations and could reduce availability of calcium, magnesium and zinc if fed in excess of the approved rate of 2% of total feed.
1990 Poultry Science 69:1364-1370
Hydrated Sodium Calcium Aluminosilicate: Effects on Zinc, Manganese, Vitamin A, and Riboflavin Utilization
Chung TK, Ekdman, Jr JW, Baker DH
"zeolite is a framework of alumino-silicate tetrahedra whose structure contains channels (known as molecular sieves in synthetic zeolites) filled with water and exchangeable cations (Mumpton and Fishman, 1977). When channels are devoid of water, adsorption properties result. "..the adsorption and ion-exchange properties of zeolites create potential for compromising mineral nutriture."
Hydrated sodium calcium aluminosilicate (HSCAS) was found to decrease zinc utilization.
According to Dr Eleanor Kellon (The Horse's Mouth), Zeolite can absorb aflatoxin but not other toxins. It can absorb dissolved positively charged molecular minerals, but it will bind nutritionally important minerals as well as toxins. It is not absorbed into the body as zeolite and has no ability to remove anything from the body through the kidneys/urine. It can block absorption in the intestines.
Zeolite is hydrated alkali aluminum silicate.
Zeolite is toxic when inhaled.
Zeolite - www.mskcc.org
NRC Nutrient Requirements of Horses 2007 p 186:
Zeolites are potential binders of cations and could reduce availability of calcium, magnesium and zinc if fed in excess of the approved rate of 2% of total feed.
1990 Poultry Science 69:1364-1370
Hydrated Sodium Calcium Aluminosilicate: Effects on Zinc, Manganese, Vitamin A, and Riboflavin Utilization
Chung TK, Ekdman, Jr JW, Baker DH
"zeolite is a framework of alumino-silicate tetrahedra whose structure contains channels (known as molecular sieves in synthetic zeolites) filled with water and exchangeable cations (Mumpton and Fishman, 1977). When channels are devoid of water, adsorption properties result. "..the adsorption and ion-exchange properties of zeolites create potential for compromising mineral nutriture."
Hydrated sodium calcium aluminosilicate (HSCAS) was found to decrease zinc utilization.
According to Dr Eleanor Kellon (The Horse's Mouth), Zeolite can absorb aflatoxin but not other toxins. It can absorb dissolved positively charged molecular minerals, but it will bind nutritionally important minerals as well as toxins. It is not absorbed into the body as zeolite and has no ability to remove anything from the body through the kidneys/urine. It can block absorption in the intestines.
Zinc
Zinc is an essential component of many enzymes.
The ratio of zinc to copper is important, and should be between 3-5 zinc:1 copper, ideally 3.5-4.5:1.
The NRC RDA for a 500 kg horse in light work or maintenance is 400 mg zinc per day.
The NRC proposed upper safe limit (for a 500 kg horse) is 500 ppm (same as mg/kg) zinc, although no toxicity was reported at 750 ppm - so for a 500 kg horse eating 2% bodyweight (10 kg DM) the upper limit for zinc would be 5000 mg/day.
Sources of Zinc
Horse feeds typically contain around 15 to 40 mg of zinc per kg dry matter (DM).
Zinc can be supplemented as:
zinc sulphate
zinc oxide
zinc chloride
zinc carbonate
organic zinc chelates
Zinc is an essential component of many enzymes.
The ratio of zinc to copper is important, and should be between 3-5 zinc:1 copper, ideally 3.5-4.5:1.
The NRC RDA for a 500 kg horse in light work or maintenance is 400 mg zinc per day.
The NRC proposed upper safe limit (for a 500 kg horse) is 500 ppm (same as mg/kg) zinc, although no toxicity was reported at 750 ppm - so for a 500 kg horse eating 2% bodyweight (10 kg DM) the upper limit for zinc would be 5000 mg/day.
Sources of Zinc
Horse feeds typically contain around 15 to 40 mg of zinc per kg dry matter (DM).
Zinc can be supplemented as:
zinc sulphate
zinc oxide
zinc chloride
zinc carbonate
organic zinc chelates
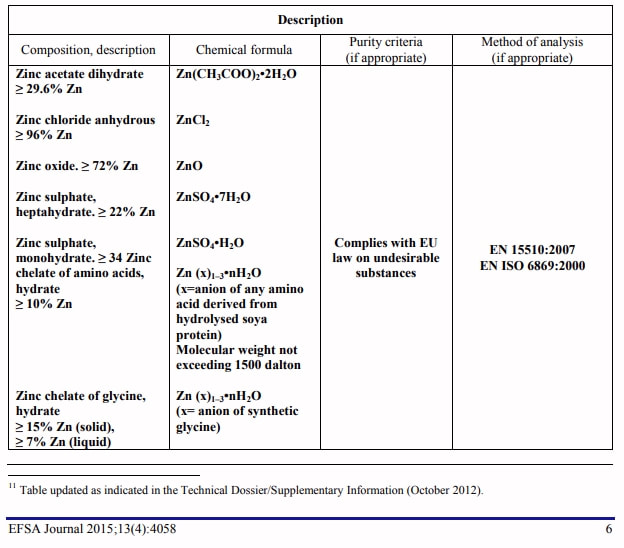
Above: Image from EFSA Scientific Opinion on the safety and efficacy of zinc compounds (E6) as feed additives for all animal species April 2016.
Research has suggested no significant difference in absorption between inorganic and organic sources of zinc. Wagner et al. 2005 found the absorption of zinc oxide to be 13.9%, zinc sulphate 12.8% and a zinc chelate 10.6% in adult horses. Baker et al. 2005 found horses retained more zinc when fed zinc oxide than a zinc chelate. Miller et al. 2003 found yearlings retained more zinc when fed organic zinc compared to inorganic zinc. Wichert et al. 2002 found higher bioavailability of zinc sulphate and zinc sulphate chelate than zinc oxide. See NRC Nutrient Requirements of Horses 2007 for details.
EFSA Scientific Opinion on the safety and efficacy of zinc compounds (E6) as feed additives for all animal species (zinc acetate, dihydrate; zinc chloride, anhydrous; zinc oxide; zinc sulphate, heptahydrate; zinc sulphate, monohydrate; zinc chelate of amino acids, hydrate; zinc chelate of glycine, hydrate) April 2016.
Zinc methionine appears to be zinc methionine sulphate
The FDA defines zinc methionine sulfate as being:
the product of the reaction between equimolar amounts of zinc sulfate and DL-methionine in purified water.
Zinc content: 19 to 22 %, DL-methionine content 46 to 50 %
EFSA: "methionine-zinc, technically pure (Met-Zinc) is zinc chelated with methionine in a molar ratio 1:2".
Minimum of 78.0 % DL-Methionine and min 17.5% Zinc and max. 18.5 % Zinc."The minimum methionine and zinc contents of the additive should be aligned with the levels set in Commission Directive 88/485/EEC: minimum 80 % methionine and maximum 18.5 % zinc".
"The FEEDAP Panel concludes that additive methionine-zinc (Met-Zinc) is safe for all animal species/categories considering that its use in supplementing feed is first limited by the regulatory maximum content of zinc. However, its contribution to dietary methionine needs
consideration when formulating diets".
Chien XX, Zafra-Stone S, Bagchi M, Bagchi D.
Bioavailability, antioxidant and immune-enhancing properties of zinc methionine.
Biofactors. 2006;27(1-4):231-44. doi: 10.1002/biof.5520270120. PMID: 17012778. (Full paper ResearchGate)
Wagner EL, Potter GD, Gibbs PG, Eller EM, Scott BD, Vogelsang MM, Walzem RL
Copper and zinc balance in exercising horses fed 2 forms of mineral supplements
J Anim Sci. 2011 Mar;89(3):722-8. doi: 10.2527/jas.2010-2871. Epub 2010 Nov 12
Studies comparing the absorption and retention of various forms of trace minerals in horses have yielded mixed results. The objective of this study was to compare Cu and Zn absorption and retention in exercising horses where the mineral was supplemented in the sulfate or organic chelate form. Nine mature horses were used in a modified switchback design experiment consisting of seven 28-d periods. Horses were fed a diet consisting of 50% concentrate and 50% hay that was balanced to meet the energy, protein, Ca, and P requirements for horses performing moderate-intensity exercise. Horses were subjected to a controlled mineral repletion-depletion diet sequence before feeding the experimental diet to standardize mineral status across horses. The experimental diet was designed to provide 90% of the 1989 NRC for Cu and Zn, with supplemental mineral provided in the inorganic sulfate form (CuSO(4) and ZnSO(4)) or the organic chelate form (Cu-Lys and Zn-Met). Feed, fecal, urine, and water samples collected during a total collection during the last 4 d of the experimental diet periods were analyzed to determine apparent absorption and retention of Cu and Zn from the 2 mineral forms. A formulation error caused horses receiving the organic chelate diet to consume about 3 times the amount of Cu and Zn compared with those fed the sulfate-supplemented diet. Copper and Zn intake and fecal excretion were greater (P < 0.05) for horses consuming the organic chelate-supplemented diet. Apparent absorption values for all horses were negative. Apparent Cu absorption and retention as a percentage of intake were greater for horses fed the organic chelate diet (P < 0.05). It is unknown why excretion of Cu and Zn by the horses during the total collection exceeded the mineral intake. Although Cu-Lys seemed to be better absorbed than CuSO(4) and absorption of Zn-Met and ZnSO(4) were not different, these results are tempered by the observation of abnormally high fecal and urinary excretion values for Cu and Zn in the present study.
Deficiency, excess, and benefits of zinc
Maximum Nutrient Values in Horse Feeds - Animal Nutrition Association of Canada August 2018
Paßlack N, van Bömmel-Wegmann S, Vahjen W, Zentek J
Impact of dietary zinc chloride hydroxide and zinc methionine on the faecal microbiota of healthy adult horses and ponies
Journal of Equine Veterinary Science published online 06 November 2021 103804. https://doi.org/10.1016/j.jevs.2021.103804
(Notes based on the abstract) 8 healthy ponies and 2 healthy horses had zinc supplemented at
1. maintenance levels 4 mg/kg BW0.75/day = 423 mg for a 500 kg horse
2. 120 mg/kg DM/day = 1200 mg for a 500 kg horse eating 2% BW per day
3. 240 mg/kg DM/day = 2400 mg for a 500 kg horse eating 2% BW per day
with either zinc chloride hydroxide or zinc methionine (6 treatments in total), each for a period of 4 weeks.
Increasing the amount of zinc reduced the richness of the faecal microbiota.
At the high zinc level, the relative abundance of Bacteroidetes and Fibrobacteres, and the amounts of acetate and total volatile fatty acids in the faeces decreased. Some effects on the microbiota were more pronounced when zinc chloride hydroxide was fed, suggesting differences in the bioavailability of organic v inorganic zinc sources.
TLS comment: other research has found that increasing the amount of zinc in the diet causes the amount of zinc in the faeces to increase. It would be interesting to know the apparent and true absorption of zinc from both sources and at all feeding rates.
Can diet prevent scratches in horses? Clair Thunes www.thehorse.com Dec 2018
"if a horse’s diet is zinc-deficient, he might not have optimal basal keratinocytes stimulation, tissue healing abilities could be reduced, and an effective barrier might not be maintained."
Agricultural & Veterinary Sciences Vol.2, No.1, 2018, pp.17-23
EFFECT OF ZINC ON INTEGRITY OF HORSE HOOF
Zuhair Noormohammady, Muhammad Chamani, Hamid Reza Khodae
Trace Mineral Basics: Zinc and Copper - www.thehorse.com July 2018
Gordon ME, Edwards MS, Sweeney CR, Jerina ML
Effects of added chelated trace minerals, organic selenium, yeast culture, direct-fed microbials, and Yucca schidigera extract in horses. Part I: Blood nutrient concentration and digestibility
Journal of Animal Science, Volume 91, Issue 8, August 2013, Pages 3899–3908, https://doi.org/10.2527/jas.2013-6122
An Evaluation of the Clinical Effect of Oral Zinc Supplementation to Horses with Distal Limb Skin Disease and an Assessment of Different Paraclinical Techniques to Estimate Equine Zinc Status
2010 Graduation research article for the completion of a Danish Certificate in Equine Medicine
Krzysztof Marycz, Eberhard Moll, Wojciech Zawadzki, Jakub Nicpoń
The Correlation of Elemental Composition and Morphological Properties of the Horses Hair after 100 Days of Feeding with High Quality Commercial Food Enriched with Zn and Cu Organic Forms 2009 Vol 12 3
Zinc deficiency may reduce thyroid hormones:Endocr Res. 2003 Nov;29(4):473-81
Pinealectomy and zinc deficiency have opposite effects on thyroid hormones in rats
Baltaci AK, Mogulkoc R, Bediz CS, Kul A, Ugur A
Research in rats showed that "free T3, T4, and TSH levels in the zinc-deficient group (Group 2) were lower than all others (p < 0.01)." "Zinc deficiency has a suppressing effect on thyroid hormones".
Higami A
Occurrence of white line disease in performance horses fed on low-zinc and low-copper diets
J Equine Science Vol, 10 No. 1 pp 1-5 1999
"the present results strongly suggested that long-term, not severe low-zinc and low-copper diets may cause a high risk of WLD in adult horses, and that an improved diet of these minerals may largely lead to the prevention of WLD."
Further info/research
Scientific Opinion on the safety and efficacy of zinc compounds (E6) as feed additives for all animal species - EFSA 2015
Osredkar J, Sustar N
Copper and Zinc, Biological Role and Significance of Copper/Zinc Imbalance
J Clinic Toxicol S3:001 2011
Zinc in the soil
Advanced Soil Geochemical Atlas of England and Wales - UK Soil Observatory
Copper and zinc content in the agricultural land of Europe - Chemistry of Europe’s Agricultural Soils. Part A 2014
Zinc in Ireland - Teagasc Agriculture and Food Development Authority
Zinc in the environment
FOREGS European map of zinc in top soil
Pollutants, Human Health and the Environment: A Risk Based Approach - Jane A. Plant, Nikolaos Voulvoulis, K. Vala Ragnar
p 66
Zinc transporters
Espinoza A, Le Blanc S, Olivares M, Pizarro F, Ruz M, Arredondo M
Iron, copper, and zinc transport: inhibition of divalent metal transporter 1 (DMT1) and human copper transporter 1 (hCTR1) by shRNA
Biol Trace Elem Res. 2012 May;146(2):281-6
Hill GM, Link JE
Transporters in the absorption and utilization of zinc and copper
J ANIM SCI April 2009 vol. 87 no. 14 suppl E85-E89
Research has suggested no significant difference in absorption between inorganic and organic sources of zinc. Wagner et al. 2005 found the absorption of zinc oxide to be 13.9%, zinc sulphate 12.8% and a zinc chelate 10.6% in adult horses. Baker et al. 2005 found horses retained more zinc when fed zinc oxide than a zinc chelate. Miller et al. 2003 found yearlings retained more zinc when fed organic zinc compared to inorganic zinc. Wichert et al. 2002 found higher bioavailability of zinc sulphate and zinc sulphate chelate than zinc oxide. See NRC Nutrient Requirements of Horses 2007 for details.
EFSA Scientific Opinion on the safety and efficacy of zinc compounds (E6) as feed additives for all animal species (zinc acetate, dihydrate; zinc chloride, anhydrous; zinc oxide; zinc sulphate, heptahydrate; zinc sulphate, monohydrate; zinc chelate of amino acids, hydrate; zinc chelate of glycine, hydrate) April 2016.
Zinc methionine appears to be zinc methionine sulphate
The FDA defines zinc methionine sulfate as being:
the product of the reaction between equimolar amounts of zinc sulfate and DL-methionine in purified water.
Zinc content: 19 to 22 %, DL-methionine content 46 to 50 %
EFSA: "methionine-zinc, technically pure (Met-Zinc) is zinc chelated with methionine in a molar ratio 1:2".
Minimum of 78.0 % DL-Methionine and min 17.5% Zinc and max. 18.5 % Zinc."The minimum methionine and zinc contents of the additive should be aligned with the levels set in Commission Directive 88/485/EEC: minimum 80 % methionine and maximum 18.5 % zinc".
"The FEEDAP Panel concludes that additive methionine-zinc (Met-Zinc) is safe for all animal species/categories considering that its use in supplementing feed is first limited by the regulatory maximum content of zinc. However, its contribution to dietary methionine needs
consideration when formulating diets".
Chien XX, Zafra-Stone S, Bagchi M, Bagchi D.
Bioavailability, antioxidant and immune-enhancing properties of zinc methionine.
Biofactors. 2006;27(1-4):231-44. doi: 10.1002/biof.5520270120. PMID: 17012778. (Full paper ResearchGate)
Wagner EL, Potter GD, Gibbs PG, Eller EM, Scott BD, Vogelsang MM, Walzem RL
Copper and zinc balance in exercising horses fed 2 forms of mineral supplements
J Anim Sci. 2011 Mar;89(3):722-8. doi: 10.2527/jas.2010-2871. Epub 2010 Nov 12
Studies comparing the absorption and retention of various forms of trace minerals in horses have yielded mixed results. The objective of this study was to compare Cu and Zn absorption and retention in exercising horses where the mineral was supplemented in the sulfate or organic chelate form. Nine mature horses were used in a modified switchback design experiment consisting of seven 28-d periods. Horses were fed a diet consisting of 50% concentrate and 50% hay that was balanced to meet the energy, protein, Ca, and P requirements for horses performing moderate-intensity exercise. Horses were subjected to a controlled mineral repletion-depletion diet sequence before feeding the experimental diet to standardize mineral status across horses. The experimental diet was designed to provide 90% of the 1989 NRC for Cu and Zn, with supplemental mineral provided in the inorganic sulfate form (CuSO(4) and ZnSO(4)) or the organic chelate form (Cu-Lys and Zn-Met). Feed, fecal, urine, and water samples collected during a total collection during the last 4 d of the experimental diet periods were analyzed to determine apparent absorption and retention of Cu and Zn from the 2 mineral forms. A formulation error caused horses receiving the organic chelate diet to consume about 3 times the amount of Cu and Zn compared with those fed the sulfate-supplemented diet. Copper and Zn intake and fecal excretion were greater (P < 0.05) for horses consuming the organic chelate-supplemented diet. Apparent absorption values for all horses were negative. Apparent Cu absorption and retention as a percentage of intake were greater for horses fed the organic chelate diet (P < 0.05). It is unknown why excretion of Cu and Zn by the horses during the total collection exceeded the mineral intake. Although Cu-Lys seemed to be better absorbed than CuSO(4) and absorption of Zn-Met and ZnSO(4) were not different, these results are tempered by the observation of abnormally high fecal and urinary excretion values for Cu and Zn in the present study.
Deficiency, excess, and benefits of zinc
Maximum Nutrient Values in Horse Feeds - Animal Nutrition Association of Canada August 2018
Paßlack N, van Bömmel-Wegmann S, Vahjen W, Zentek J
Impact of dietary zinc chloride hydroxide and zinc methionine on the faecal microbiota of healthy adult horses and ponies
Journal of Equine Veterinary Science published online 06 November 2021 103804. https://doi.org/10.1016/j.jevs.2021.103804
(Notes based on the abstract) 8 healthy ponies and 2 healthy horses had zinc supplemented at
1. maintenance levels 4 mg/kg BW0.75/day = 423 mg for a 500 kg horse
2. 120 mg/kg DM/day = 1200 mg for a 500 kg horse eating 2% BW per day
3. 240 mg/kg DM/day = 2400 mg for a 500 kg horse eating 2% BW per day
with either zinc chloride hydroxide or zinc methionine (6 treatments in total), each for a period of 4 weeks.
Increasing the amount of zinc reduced the richness of the faecal microbiota.
At the high zinc level, the relative abundance of Bacteroidetes and Fibrobacteres, and the amounts of acetate and total volatile fatty acids in the faeces decreased. Some effects on the microbiota were more pronounced when zinc chloride hydroxide was fed, suggesting differences in the bioavailability of organic v inorganic zinc sources.
TLS comment: other research has found that increasing the amount of zinc in the diet causes the amount of zinc in the faeces to increase. It would be interesting to know the apparent and true absorption of zinc from both sources and at all feeding rates.
Can diet prevent scratches in horses? Clair Thunes www.thehorse.com Dec 2018
"if a horse’s diet is zinc-deficient, he might not have optimal basal keratinocytes stimulation, tissue healing abilities could be reduced, and an effective barrier might not be maintained."
Agricultural & Veterinary Sciences Vol.2, No.1, 2018, pp.17-23
EFFECT OF ZINC ON INTEGRITY OF HORSE HOOF
Zuhair Noormohammady, Muhammad Chamani, Hamid Reza Khodae
Trace Mineral Basics: Zinc and Copper - www.thehorse.com July 2018
Gordon ME, Edwards MS, Sweeney CR, Jerina ML
Effects of added chelated trace minerals, organic selenium, yeast culture, direct-fed microbials, and Yucca schidigera extract in horses. Part I: Blood nutrient concentration and digestibility
Journal of Animal Science, Volume 91, Issue 8, August 2013, Pages 3899–3908, https://doi.org/10.2527/jas.2013-6122
An Evaluation of the Clinical Effect of Oral Zinc Supplementation to Horses with Distal Limb Skin Disease and an Assessment of Different Paraclinical Techniques to Estimate Equine Zinc Status
2010 Graduation research article for the completion of a Danish Certificate in Equine Medicine
Krzysztof Marycz, Eberhard Moll, Wojciech Zawadzki, Jakub Nicpoń
The Correlation of Elemental Composition and Morphological Properties of the Horses Hair after 100 Days of Feeding with High Quality Commercial Food Enriched with Zn and Cu Organic Forms 2009 Vol 12 3
Zinc deficiency may reduce thyroid hormones:Endocr Res. 2003 Nov;29(4):473-81
Pinealectomy and zinc deficiency have opposite effects on thyroid hormones in rats
Baltaci AK, Mogulkoc R, Bediz CS, Kul A, Ugur A
Research in rats showed that "free T3, T4, and TSH levels in the zinc-deficient group (Group 2) were lower than all others (p < 0.01)." "Zinc deficiency has a suppressing effect on thyroid hormones".
Higami A
Occurrence of white line disease in performance horses fed on low-zinc and low-copper diets
J Equine Science Vol, 10 No. 1 pp 1-5 1999
"the present results strongly suggested that long-term, not severe low-zinc and low-copper diets may cause a high risk of WLD in adult horses, and that an improved diet of these minerals may largely lead to the prevention of WLD."
Further info/research
Scientific Opinion on the safety and efficacy of zinc compounds (E6) as feed additives for all animal species - EFSA 2015
Osredkar J, Sustar N
Copper and Zinc, Biological Role and Significance of Copper/Zinc Imbalance
J Clinic Toxicol S3:001 2011
Zinc in the soil
Advanced Soil Geochemical Atlas of England and Wales - UK Soil Observatory
Copper and zinc content in the agricultural land of Europe - Chemistry of Europe’s Agricultural Soils. Part A 2014
Zinc in Ireland - Teagasc Agriculture and Food Development Authority
Zinc in the environment
FOREGS European map of zinc in top soil
Pollutants, Human Health and the Environment: A Risk Based Approach - Jane A. Plant, Nikolaos Voulvoulis, K. Vala Ragnar
p 66
Zinc transporters
Espinoza A, Le Blanc S, Olivares M, Pizarro F, Ruz M, Arredondo M
Iron, copper, and zinc transport: inhibition of divalent metal transporter 1 (DMT1) and human copper transporter 1 (hCTR1) by shRNA
Biol Trace Elem Res. 2012 May;146(2):281-6
Hill GM, Link JE
Transporters in the absorption and utilization of zinc and copper
J ANIM SCI April 2009 vol. 87 no. 14 suppl E85-E89